
PHERAstar FSX
Powerful and most sensitive HTS plate reader


Barry Whyte is Application Scientist and Science Writer at BMG LABTECH in the United States. He has PhD and Bachelor of Science (BSc) degrees in biochemistry from the University of Bristol in the United Kingdom and more than 20 years of experience in the life sciences and science communications. Over the years, Barry has worked on three continents and traveled widely. He enjoys building on his international work experience and learning new ways to help scientists advance their research.
While metabolism is a universal attribute of all cells, no one-fit playbook exists for the types of metabolism present and operating in different types of cells. Cancer cells are no exception.
Different cancer cells show significant changes in metabolism compared with healthy cells. Amongst the most common alterations found in various cancer cells are increased glucose uptake and lactate production, which are hallmarks of metabolism in cancer cells, dependencies on amino acids like glutamine or proline, enhanced lipid synthesis, and changes in mitochondrial function. In cancer cells, major changes in gene expression help increase glucose uptake to support their rapid growth. It is known that most cancer cells rely on glycolysis to drive cellular processes even when oxygen is available unlike normal differentiated cells that primarily rely on mitochondrial oxidative phosphorylation (this is known as the Warburg effect which is discussed later in this blog). Collectively, these differences in metabolism between healthy cells and various cancer cells are often referred to as tumor metabolism, which encompasses the altered metabolic processes that support tumor growth and survival.
Researchers are still trying to untangle the complexity of these differences in the metabolism of diverse cancer cells which have repercussions not only for understanding how cells work but also for new approaches to drug development.
Knowledge of metabolism in cancer cells is built upon years of major scientific findings that have established the core metabolic concepts and pathways in operation in both microbes and eukaryotic cells. Discoveries across many disciplines including biochemistry, cell biology, and molecular biology have shaped what is known today about metabolism in the life sciences.1-3 In the context of metabolism in cancer, these foundational studies have provided critical insights into how metabolic pathways are altered in tumor cells. Before taking a closer look at metabolism in cancer, it is worth revisiting some of the tenets of regular metabolism.
Metabolism comprises the chemicals, enzymes and metabolic pathways that make life possible.3,4 Cells of all types, irrespective of their origin or whether they are unicellular, multicellular, naturally occurring or engineered rely on metabolism to support the processes of life.
While it is true that we have a good understanding of what constitutes cellular metabolism, this knowledge has arisen mainly from studies of cells that do not divide rapidly and from many of the cell types that are the bedrock of differentiated tissues.4 Proliferating cells, in contrast, have distinct metabolic requirements compared to non-dividing cells, as they activate pathways such as glycolysis to support rapid growth and division.
Much therefore remains to be discovered including the precise details of the subtle metabolic differences that take place in different cell types, various disease states like cancer, or under different growth conditions. Additionally, limitations on the endogenous levels of nutrients in the cell act as an internal constraint that regulates or restricts cell proliferation and metabolism in healthy cells.
Overall, the pathways of metabolism can be broadly classified into reactions that either synthesize larger molecules (anabolic reactions) or those that breakdown larger molecules (Fig.1). These two major groups either generate molecules that are useful to build the larger more complex molecules, which are essential for the operation and survival of the cell, or liberate the energy that can be used by the cell to run essential cellular processes.3 These processes are vital for life. They can also be vital for the proliferation of cancer cells. In addition, futile metabolic cycles can occur, consuming energy without productive output, and play a significant role in metabolic regulation.
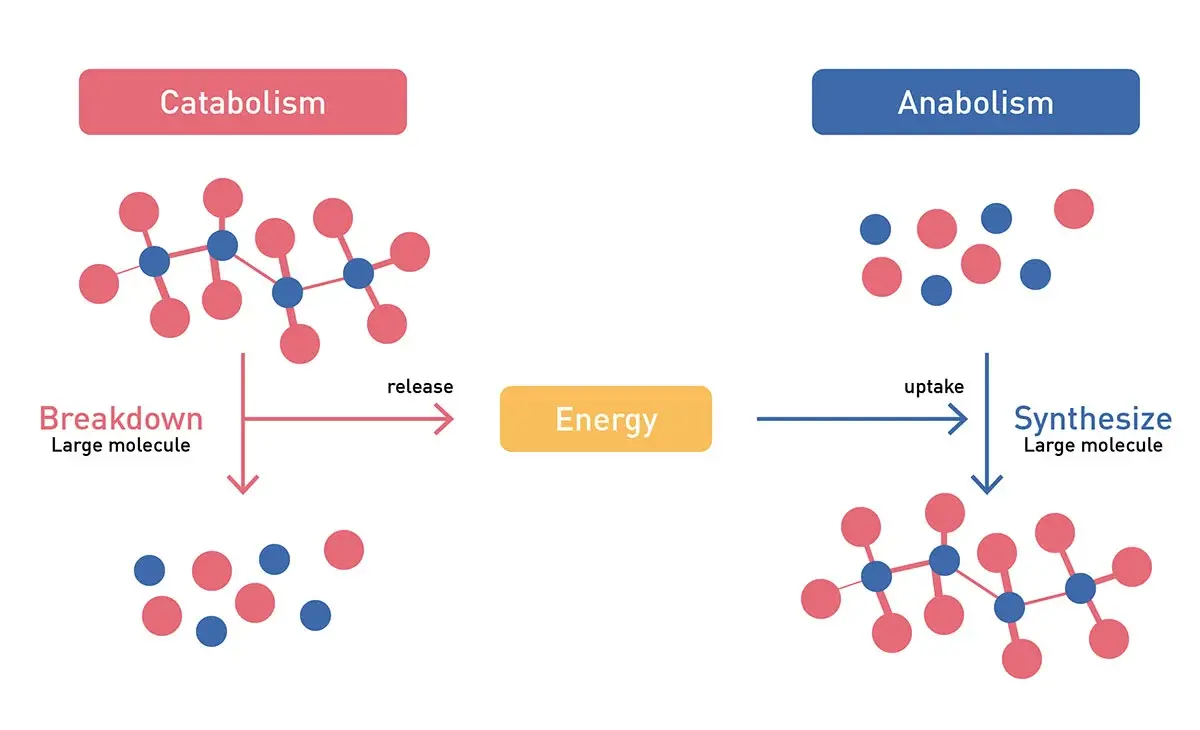
The major metabolic pathways that produce energy include glycolysis (glucose to pyruvate, ATP and NADH production), the citric acid cycle (acetyl-CoA to carbon dioxide, NADH, FADH2, and ATP production), and oxidative phosphorylation (ATP). In addition, a host of anabolic and biosynthetic pathways give rise to the amino acids (e.g. glutamine, alanine), nucleotides (e.g. ATP, GTP, NADH, NADPH), sugars (glucose, lactose), and fatty acids (e.g. palmitic acid, stearic acid, oleic acid) essential for building and operating a cell. This includes gluconeogenesis, the pentose-phosphate pathway, and specific pathways generating fatty acids and amino acids. These pathways are discussed in more detail in the following BMG LABTECH blogs: Cell metabolism and Microbiological applications for bacterial metabolism on a microplate reader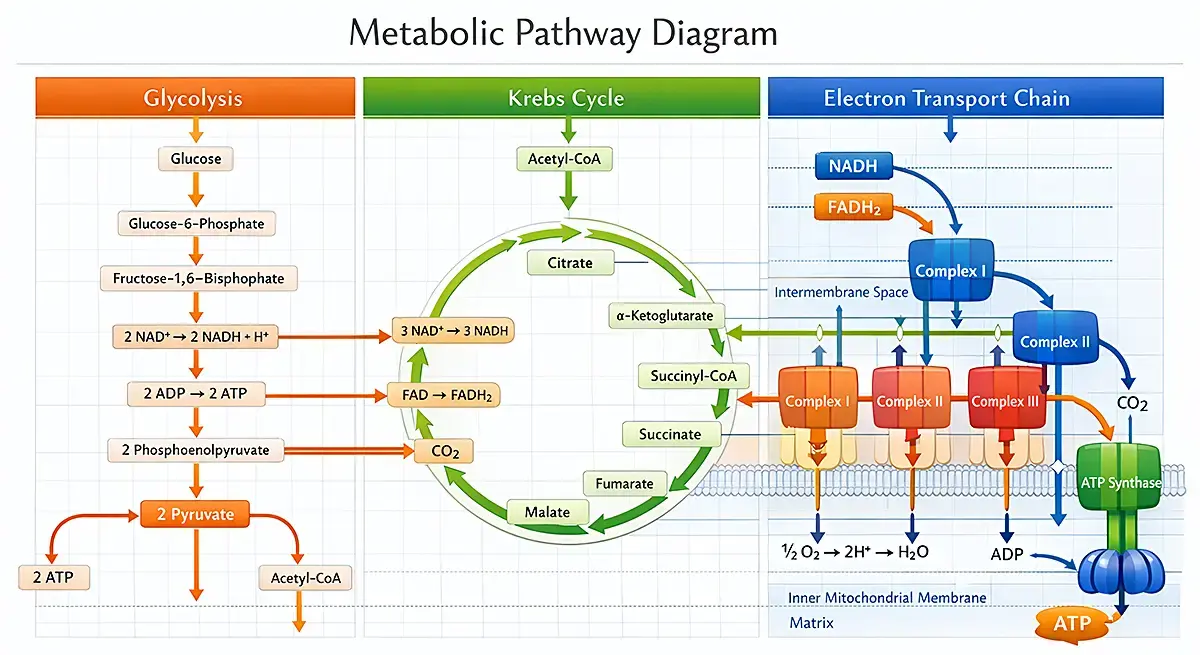
The pathways of cell metabolism are not isolated (Fig. 2) but form a highly interconnected metabolic network that spans different locations in the cell (e.g. cytoplasm, the mitochondria) and they drive different processes essential for cell viability and cell proliferation that may be involved in health or disease. Each of these metabolic pathways is also tightly linked to the regulation of the cell cycle, influencing cell division and proliferation.
Metabolism in cancer cells refers to the altered metabolic processes that occur in cancer cells. The metabolic changes observed in different cancer cells vary, reflecting significant metabolic heterogeneity among tumor types. Amongst the most common alterations are increased glucose uptake and lactate production — hallmarks of metabolism in cancer cells — dependency on amino acids like glutamine or proline, enhanced lipid synthesis, and changes in mitochondrial function.
Mitochondrial toxicity and dysfunction itself can disrupt energy production and metabolic signaling significantly, further impacting metabolism in cancer cells and contributing to tumor progression. The role of mitochondrial metabolism is therefore critical, as reprogramming of these pathways supports cancer cell survival, drug resistance, and adaptation to nutrient and oxygen limitations.
An important point also to note is that cancer cells often exhibit metabolic plasticity, allowing them to adapt to nutrient-poor or hypoxic environments by switching between glycolysis, oxidative phosphorylation, and other alternative energy sources.
Collectively, these adapted metabolic processes allow cancer cells to grow and proliferate rapidly by reprogramming normal cellular energy to support tumor growth.
One of the earliest differences noted in metabolism between normal and cancerous cells was the observation that cancer cells tend to favor glycolysis for energy production over oxidative phosphorylation even in the presence of sufficient oxygen. This observation is known as the Warburg effect after the scientist Otto Heinrich Warburg (see Fig. 3) who first made this discovery in 1923.5 The Warburg effect is characterized by the preferential conversion of glucose to lactate (lactic acid) even in the presence of oxygen and functional mitochondria. While the observations made by Otto Warburg in 1923 related to metabolic shifts in cancer still hold true, the precise details of how the Warburg effect is directly involved in cancer remain unclear.  On one level, it is thought that cancer cells prefer glycolysis because it provides not only energy but also metabolic intermediates necessary for rapid cell growth and proliferation, despite being less efficient than oxidative phosphorylation. In real terms, glycolysis provides significantly less return on energy under the same conditions when compared with oxidative phosphorylation as measured in yield of ATP (see Fig.4). However, this amount of energy is sufficient to support cell proliferation through aerobic glycolytic metabolism and is compensated by the support it offers for rapid biomass formation (nucleotides, lipids and proteins), a key driver of cell proliferation.
On one level, it is thought that cancer cells prefer glycolysis because it provides not only energy but also metabolic intermediates necessary for rapid cell growth and proliferation, despite being less efficient than oxidative phosphorylation. In real terms, glycolysis provides significantly less return on energy under the same conditions when compared with oxidative phosphorylation as measured in yield of ATP (see Fig.4). However, this amount of energy is sufficient to support cell proliferation through aerobic glycolytic metabolism and is compensated by the support it offers for rapid biomass formation (nucleotides, lipids and proteins), a key driver of cell proliferation. 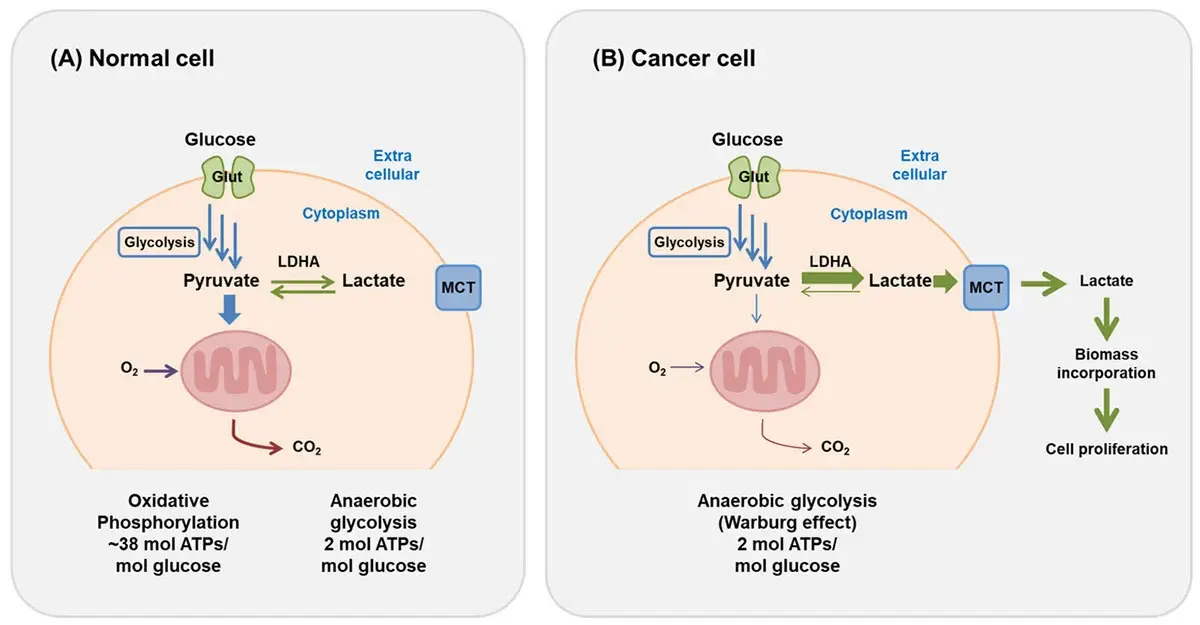 The Warburg effect is the basis for tumor detection in positron emission tomography (PET) scans, where the radioactive glucose analog [18F]fluorodeoxyglucose is taken up by cancer cells, phosphorylated by hexokinase, and trapped inside the cell, allowing visualization of tumors due to their heightened glucose uptake. PET scanning is widely used in oncology for cancer staging, therapeutic planning, and response assessment. Additionally, hyperpolarized 13C magnetic resonance imaging is a non-radioactive technique that can visualize metabolic changes in vivo and assess therapeutic effectiveness early in treatment.
The Warburg effect is the basis for tumor detection in positron emission tomography (PET) scans, where the radioactive glucose analog [18F]fluorodeoxyglucose is taken up by cancer cells, phosphorylated by hexokinase, and trapped inside the cell, allowing visualization of tumors due to their heightened glucose uptake. PET scanning is widely used in oncology for cancer staging, therapeutic planning, and response assessment. Additionally, hyperpolarized 13C magnetic resonance imaging is a non-radioactive technique that can visualize metabolic changes in vivo and assess therapeutic effectiveness early in treatment.
The presence of the Warburg effect has been described in most tumor types, including glioblastoma, pancreas, breast, and cervical tumors.
The tumor microenvironment is a dynamic and complex ecosystem that plays a pivotal role in cancer development, progression, and therapeutic response. Composed of a diverse array of cell types — including cancer cells, immune cells, stromal cells, and vascular cells — the tumor microenvironment forms a supportive network that not only nurtures tumor growth but also shapes the metabolic fate of cancer cells (Fig. 5).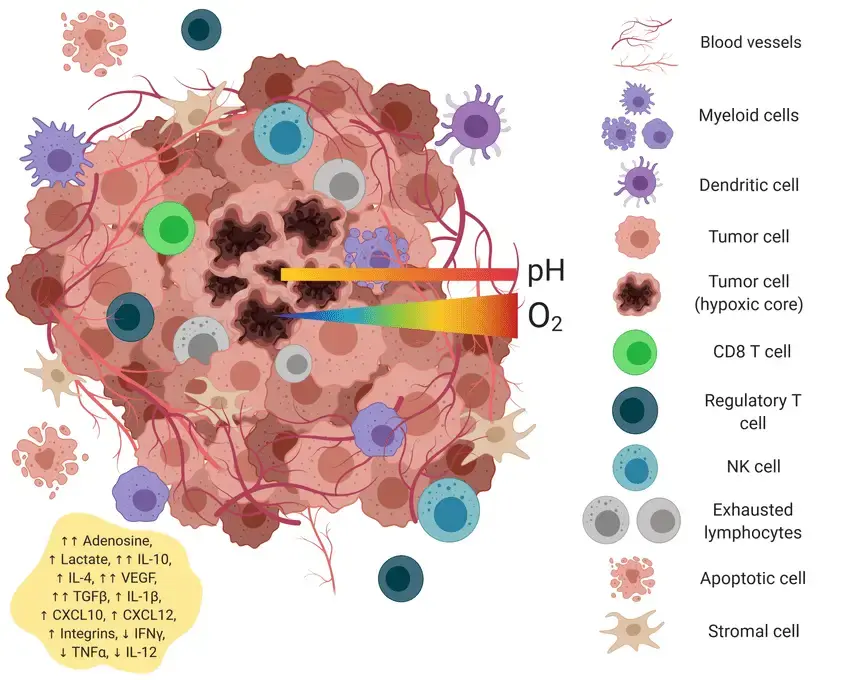 One of the defining features of the tumor microenvironment is its influence on the metabolism in cancer cells. Within this environment, cancer cells undergo profound metabolic reprogramming, including the Warburg effect we described earlier. This shift enables tumor cells to rapidly generate both energy and the molecular building blocks required for cell proliferation, supporting the aggressive growth seen in many cancers such as breast cancer and pancreatic cancer.
One of the defining features of the tumor microenvironment is its influence on the metabolism in cancer cells. Within this environment, cancer cells undergo profound metabolic reprogramming, including the Warburg effect we described earlier. This shift enables tumor cells to rapidly generate both energy and the molecular building blocks required for cell proliferation, supporting the aggressive growth seen in many cancers such as breast cancer and pancreatic cancer.
The metabolic landscape of the tumor microenvironment is further shaped by the interactions between cancer cells and surrounding stromal cells, such as cancer-associated fibroblasts. These stromal cells can secrete metabolites like lactate, which not only fuel tumor growth but also contribute to the immunosuppressive environment that allows cancer cells to evade immune surveillance. The exchange of metabolic signals —often referred to as metabolic crosstalk — between cancer cells, stromal cells, and immune cells is a key driver of tumor progression and metastasis.
Moreover, the tumor microenvironment can also modulate the expression of glycolytic enzymes and other metabolic genes in cancer cells, reinforcing their metabolic flexibility and adaptability. For example, upregulation of the pentose phosphate pathway within tumor tissues provides both reducing power and biosynthetic precursors, further supporting the needs of rapidly dividing cells.
Recent advances in cancer research have underscored the therapeutic potential of targeting these metabolic vulnerabilities. Inhibiting key steps in the glycolytic pathway or blocking lactate production and transport can disrupt the metabolic support system of tumor cells, impairing their growth and survival. Similarly, interventions aimed at the pentose phosphate pathway or other metabolic interactions within the tumor microenvironment are being explored as innovative strategies for cancer treatment.
In summary, the tumor microenvironment is not merely a passive backdrop but an active participant in cancer progression, orchestrating a network of metabolic interactions that sustain tumor growth and resistance to therapy. Understanding and targeting the unique metabolic characteristics of cancer cells within the tumor microenvironment — such as enhanced glucose metabolism, aerobic glycolysis, and metabolic crosstalk — hold significant promise for the development of more effective cancer therapies.
Evidence for differences in metabolism in normal and cancerous cells has been around for some time. However, only more recently have researchers started to look at how these differences might be channeled into the development of new drugs or cancer treatments. Current trends in cancer research are focused on understanding how metabolic pathways, cancer-related metabolites, altered metabolic flux and genetic mutations drive cancer progression and reveal new therapeutic targets. Targeting the unique dependencies of metabolism in cancer cells could lead to the development of more effective therapies with fewer side effects.
Cancer therapy strategies are increasingly focusing on targeting metabolic pathways, such as inhibiting glycolytic enzymes or exploiting metabolic vulnerabilities to disrupt cancer cell survival. Many approaches exist. Some cancers have been shown to use insulin which directly impacts metabolism as a mechanism of resistance to some potent anticancer drugs. Other cancers are addicted to amino acids like proline or glutamine.6,7 If researchers can starve cancer cells of amino acids like proline, they may be able to weaken cancer cells specifically to improve existing or new treatments. Renal cancer is one example where metabolic reprogramming and mitochondrial function play a significant role in disease progression and therapy response. Researchers are also interested in developing web-based tools to target the specific metabolic vulnerabilities of cancer cells, enabling more precise and effective therapies.
Whatever the trajectory, researchers need high quality detection methods and more innovation to delineate the intricacies of metabolism in cancer cells.
They also need robust in vivo evidence from animal model systems or clinical studies to validate these findings and therapeutic approaches related to metabolism in cancer.
Measurement of metabolism in cancer cells gives scientists a gateway to understanding many molecular processes and is important for diverse applications in the life sciences, including cell metabolism research. As we have seen earlier, these measurements can illuminate new avenues of research or help discover novel therapeutic strategies. By enabling metabolic assays in a high‑throughput format, microplate readers allow researchers to rapidly interrogate metabolic pathways, compare conditions, and uncover novel therapeutic strategies. Here we look at some of the applications of either direct or indirect detection measurements and assays and how these assays are used with microplate readers to advance scientific research.
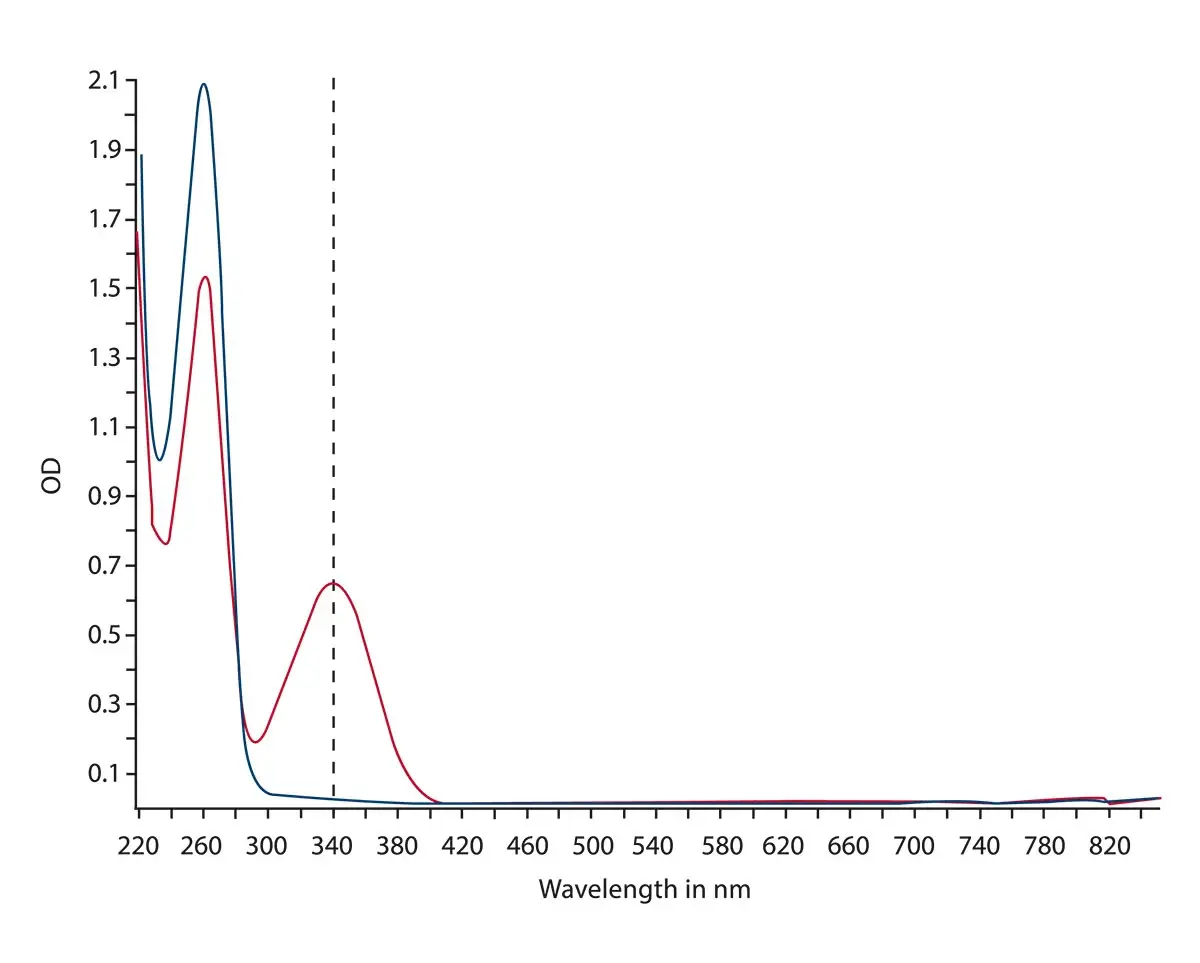
In the application note Overview of ELISA assays and NADH/NADPH conversion detection the main absorbance assays were measured using spectrometer-based microplate readers and includes NADH/NADPH conversions. NADH/NAD+ and NADPH/NADP+ are cofactors used by many enzymes in numerous cellular functions, including energy metabolism, mitochondrial function, as well as aging and cell death. Due to their direct involvement in energy metabolism, these cofactors are directly relevant to measurements of metabolism in cancer. In the case of NADH/NADPH conversions, the reaction can be monitored by measuring the absorbance at 340 nm since the oxidized forms do not absorb light at this wavelength. This can be rapidly and accurately done using the ultrafast spectrometer. The microplate readers from BMG LABTECH are all equipped with the ultrafast spectrometer that can capture the full-absorbance data at a resolution of 1 nm for all wavelengths from 220 to 1000 nm in less than one second per well. The absorbance spectra for the conversion of NAD+ to NADH are shown in Fig.6.
In the application note Glucose assay and lactate assay allow to monitor cellular glucose metabolism precisely luminescent assays for lactate and glucose were measured on the VANTAstar® microplate reader (Fig. 7). Lactate and glucose measurements are crucial for many measurements of metabolism in cancer cells. Glucose is metabolized into pyruvate in the cell. Under aerobic conditions, pyruvate is converted to acetyl-CoA which enters the citric acid cycle. When anaerobic conditions exist, pyruvate is reduced to lactate. Lactate can also originate in cancer cells from glycolysis under aerobic conditions. Measurement of lactate as well as glucose levels therefore provide valuable indicators of metabolic activity in cancer cells. Analysis of these metabolites may be needed at higher throughout for applications like drug screening and this is driving the need for microplate-based assay methods. In this context, luminescence-based assays are good rapid, highly sensitive options for researchers. In this instance, the application note mentioned above describes how to perform and optimize Lactate-GloTM and a Glucose-GloTM assays on the VANTAstar microplate reader. The Enhanced Dynamic Range of the VANTAstar microplate reader makes these assays easy to perform and eliminates the need for manual intervention from the start of the reactions to the luminescent readout. In addition, the automatic crosstalk reduction feature reduces signal interference caused by the stray light emitted by the surrounding wells of a microplate while detecting the light signal of a specific well. This allows steps to be taken by researchers to minimize crosstalk and improve performance.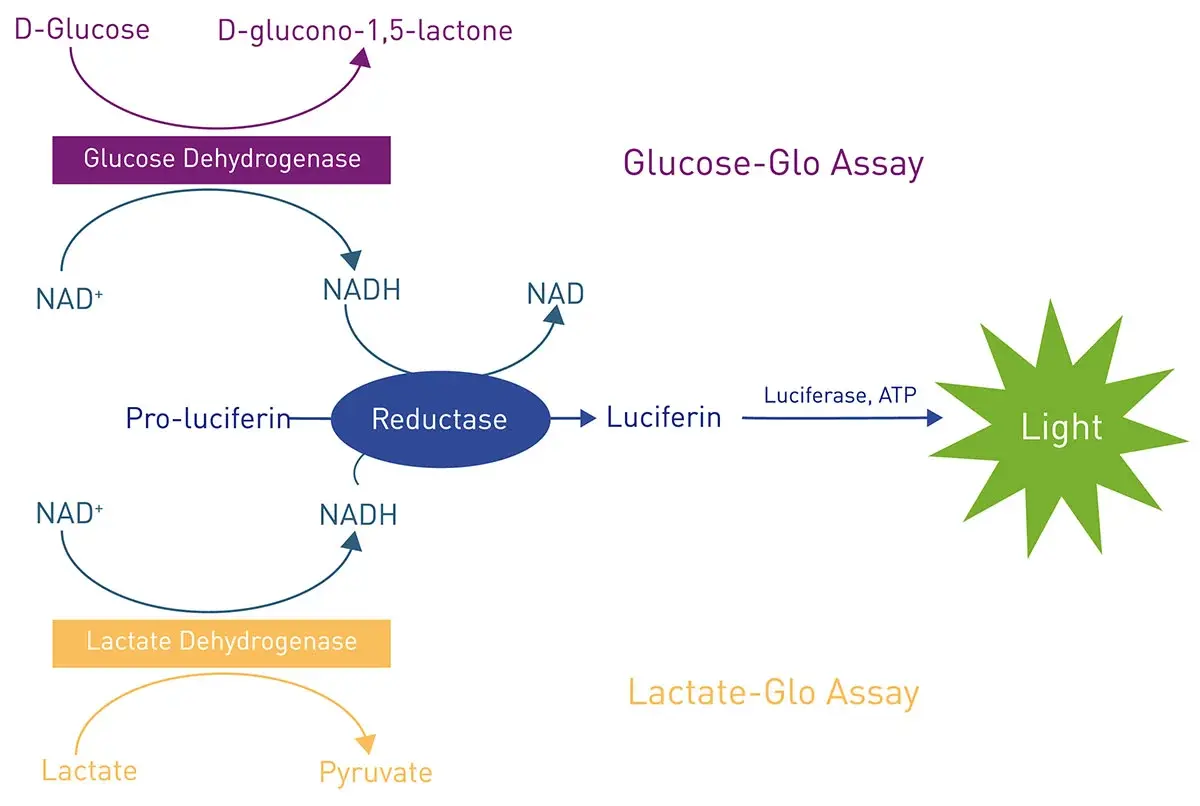
ATP and ADP are important molecules in the metabolic landscape of any cell. Many classes of enzymes, including those implicated in the metabolism of cancer, generate ADP as part of their catalytic reaction. In many cases, it is useful to have direct readouts of ADP or ADP levels in one-step homogeneous formats that can be scaled for higher throughput. The application note Transcreener ADP2 FI assay performed on BMG LABTECH microplate readers describes use of the Transcreener® technology to quantify the production of ADP during enzyme reactions (Fig.8). These assays can be performed on BMG LABTECH microplate readers with excellent Z prime values and different detection modes (fluorescence, fluorescence polarization and time-resolved fluorescence resonance energy transfer).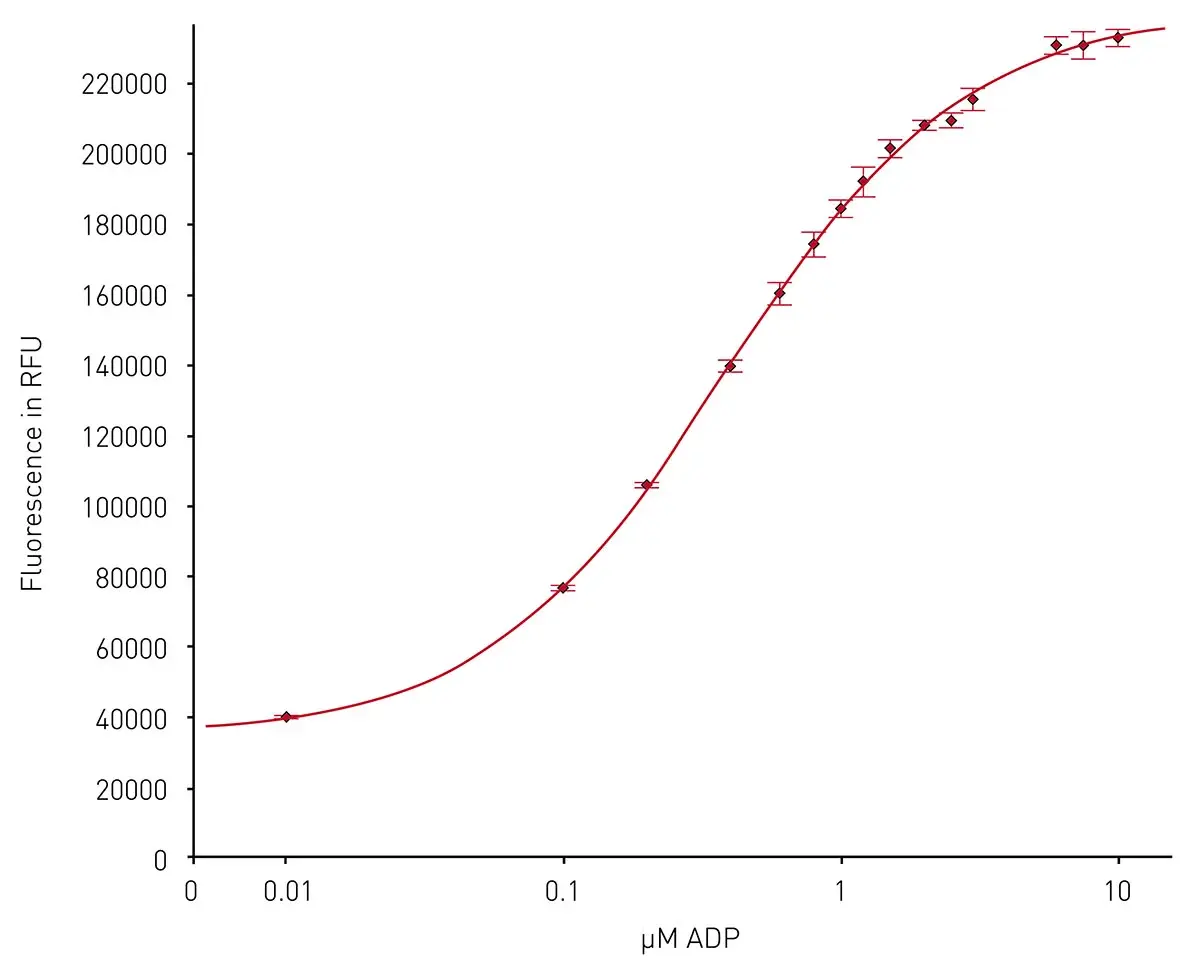
Cell viability is a crucial parameter in many disease-related studies and metabolism in cancer is no exception. Researchers are often interested in finding compounds that selectively kill cancer cells, and this includes molecules that may interfere with different metabolic steps within these cells. Toxicity studies, drug screening and basic research for metabolism in cancer cells all benefit from commercially available cell viability kits that are compatible with microplate readers.
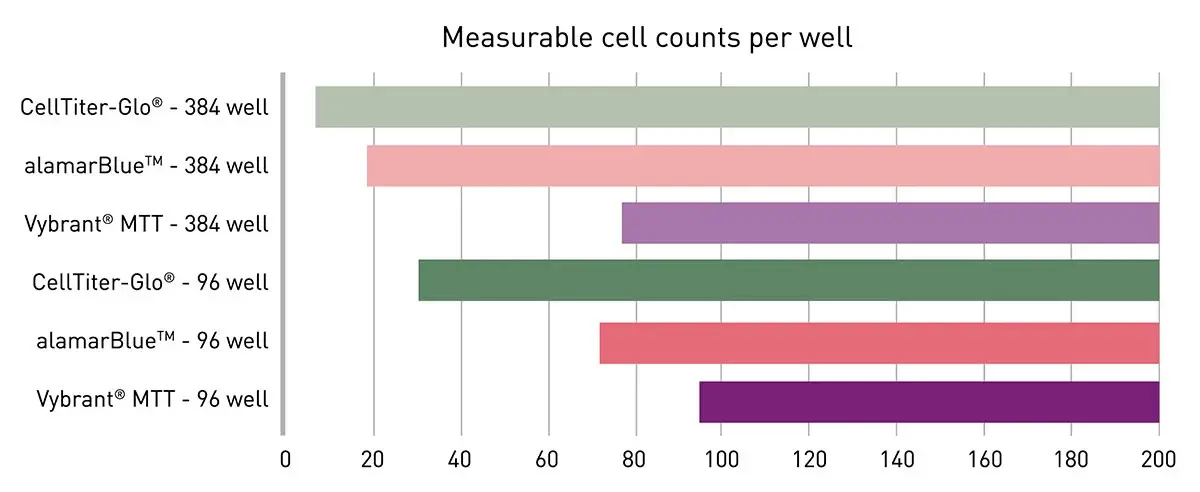
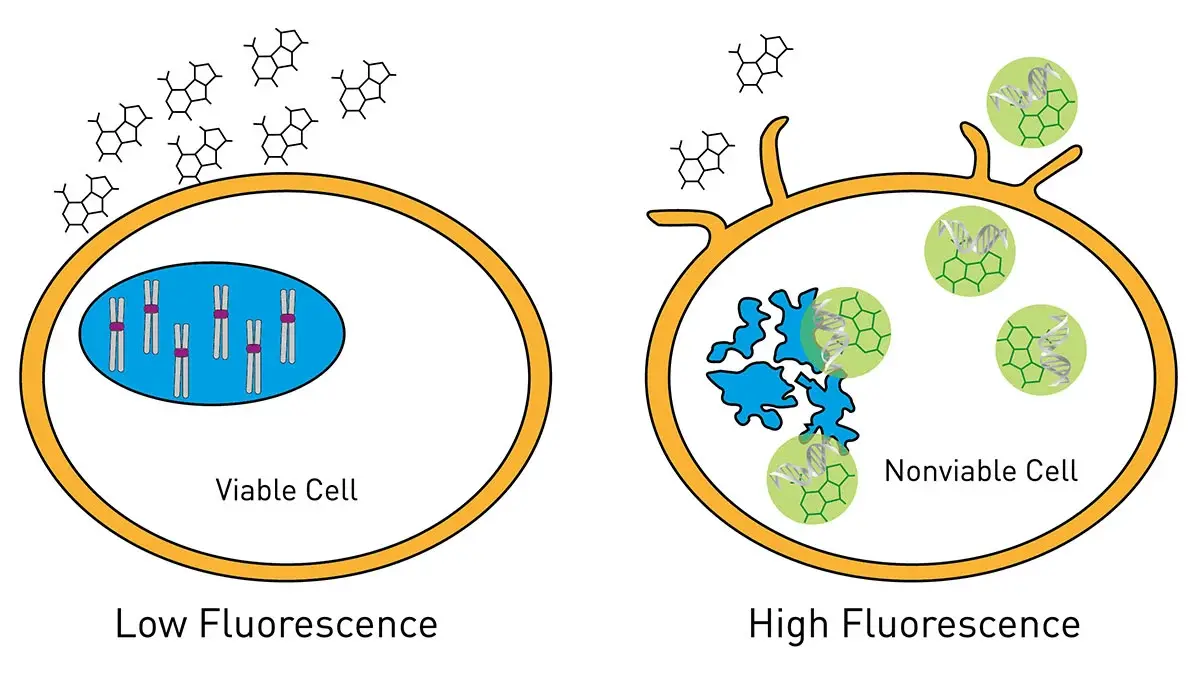
In the application note Viability assays: a comparison of luminescence-, fluorescence-, and absorbance-based assays to determine viable cell counts several options for cell viability assessment were compared for their limit of detection when used with HeLa cells, an immortal cell line originally derived in 1951 from cervical cancer cells and a potent tool for cancer research (Fig.9). The VANTAstar microplate reader provided an ideal detection platform to cover a broad range of cell viability assay read-outs including absorbance, fluorescence and luminescence measurements at high sensitivity. The high-performance LVF MonochromatorTM and spectrometer offer full flexibility in wavelength selection and accordingly in the choice of the viability assay. The use of Enhanced Dynamic Range and automatic focus adjustment eliminated the need for a preliminary gain or focus height optimization. The Vybrant® MTT, AlamarBlueTM and CellTiter-Glo® assays with colorimetric, fluorescence and luminescent readouts, respectively, were tested and offered different levels of sensitivity with outstanding performance in all conditions tested.
Cell toxicity assays are critical in studies of metabolism in cancer cells since they link metabolic data to biological viability. This helps avoid misinterpretation of metabolic readouts. They can also aid in distinguishing metabolic effects from cell death and assist in differentiating adaptive metabolism from lethal failure. The application note The new Atmospheric Control Unit (ACU) for the CLARIOstar provides versatility in long-term cell-based assays outlines how cell-based assays for cell viability and cytotoxicity can be detected in real time. For this type of measurement, it is important to keep the cells under the best possible conditions for as long as the measurements need to be made. The CLARIOstar® Plus with an Atmospheric Control Unit allowed K562 cells, a human immortalized cell line originally derived from a patient with leukemia, to be grown and tested with a panel of test compounds every hour for 72 hours. This type of incubation range is suitable for the study of many metabolic measurements in the metabolism of cancer. Real-time detection allows identification of the precise point in time when a cytotoxic or antiproliferative change occurs. The RealTime-Glo® MT Cell Viability Assay and CellTox™ Green Cytotoxicity Assay (Fig.10) were used with luminescent and fluorescent detection, respectively.
Hypoxia and subsequent reperfusion in cancer are caused by abnormal, poorly organized tumor vasculature and fluctuating blood flow, leading to repeated cycles of oxygen deprivation and reoxygenation that drive metabolic reprogramming, oxidative stress, and tumor progression. The possibility to mimic conditions where oxygen levels vary is directly useful for studies of tumor development where important changes may also be taking place in metabolic events (we mentioned the tumor microenvironment earlier). Low oxygen for example stabilizes hypoxia-inducible factor (HIF) transcription factors, promoting glycolysis, limiting mitochondrial respiration, and altering angiogenesis and nutrient use. In the application note The CLARIOstar with ACU exposes cells to ischemia-reperfusion conditions and monitors their oxygenation the ability to modulate oxygen levels over a period of hours is demonstrated in metabolically active cells. This type of experimental set up to mimic hypoxic conditions is extremely useful to probe metabolism in cancer cells since it reflects the real type of environmental conditions that cancer cells may face in the body (Fig. 11).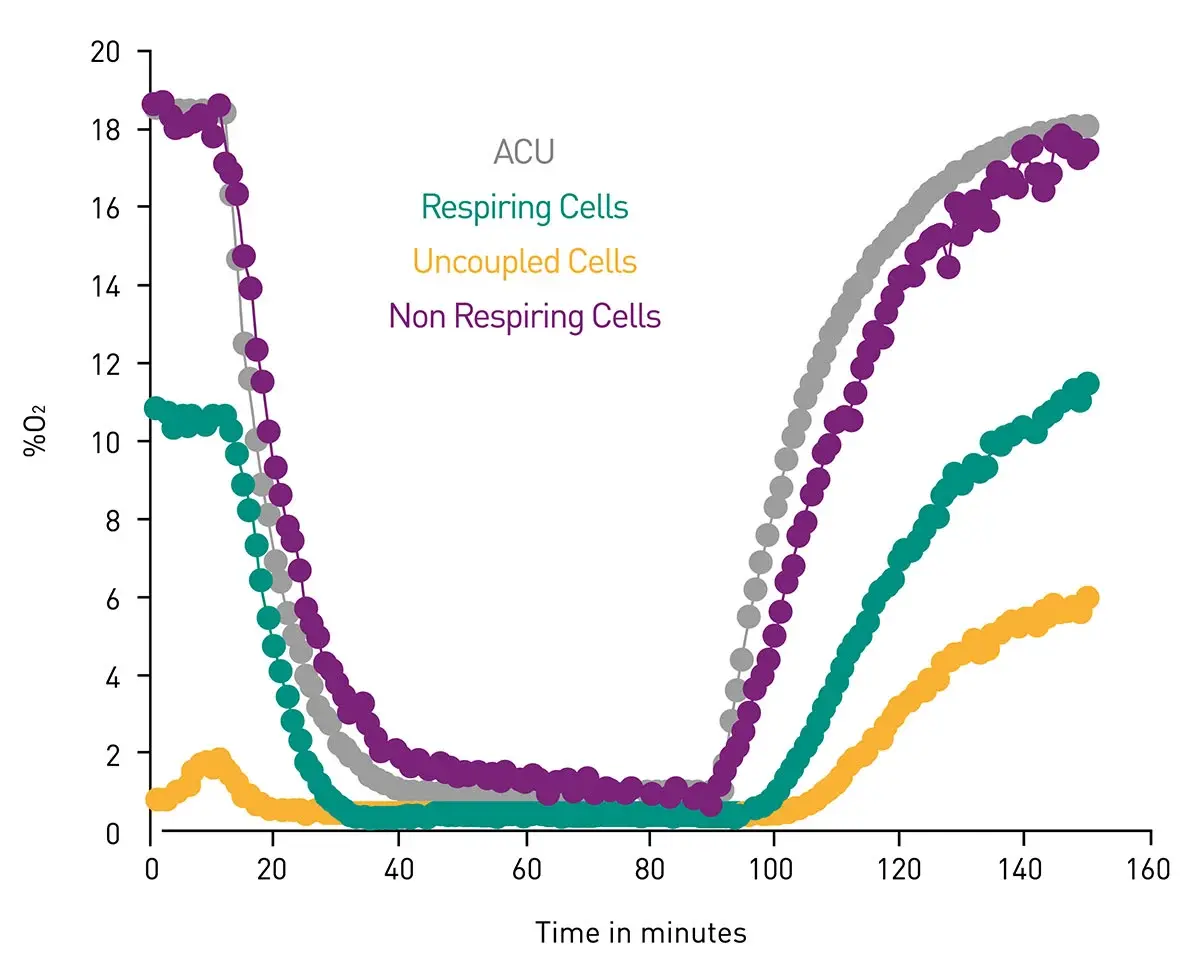
Extracellular acidification and oxygen consumption are further useful endpoints for the study of metabolism including investigations of metabolism in cancer cells. In the application note Measuring changes in cellular metabolism by monitoring extracellular acidification and oxygen consumption in real-time oxygen and pH levels were measured in real time under physiologically relevant atmospheric conditions. The system was developed on a CLARIOstar microplate reader equipped with an atmospheric control unit. The Atmospheric Control Unit uniquely enables control of both the oxygen and carbon dioxide concentrations in an independent manner. Advanced phosphorescent dyes were used to measure O2 consumption and glycolytic flux in an open-flow respirometry system. The phosphorescent oxygen and pH probe (Cayman Chemical) combined with the CLARIOstar and Atmospheric Control Unit provided a robust and efficient means of measuring cellular bioenergetics at physiologically relevant oxygen concentrations. Additional utility is derived from the ability to easily use non-adherent cells and to perform additional multiplexing to examine additional cellular responses including those linked to metabolism and cancer.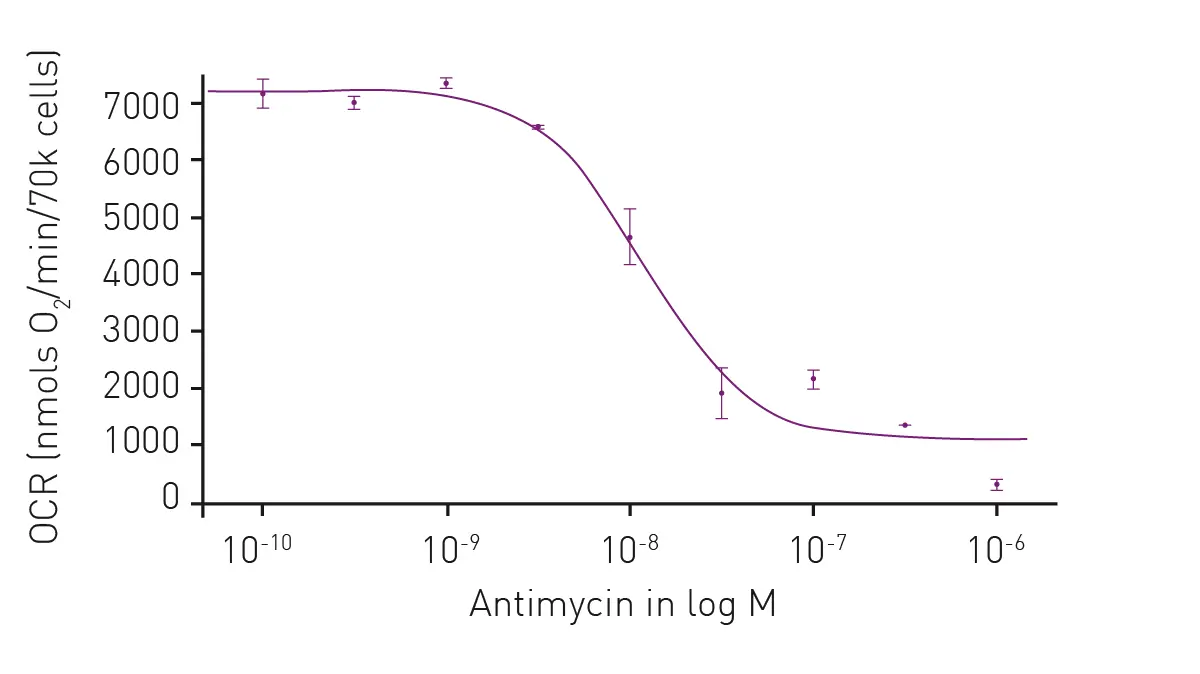
The use of gene reporter assays with fluorophores like green fluorescent protein (GFP) are also useful for making measurements related to studies of metabolism in cancer cells. In the application note Real-time monitoring of genetically encoded redox probes in mammalian cell monolayers, the green fluorescent protein roGFP2 was used as a probe to measure H2O2 levels in a monolayer of confluent lung adenocarcinoma cells. Redox processes and production of reactive oxygen species (ROS) play an important role in cellular metabolism and diseases like cancer. In particular, redox homeostasis is crucial for cancer cell survival, influencing DNA repair, tumor microenvironment modulation, and resistance to therapies such as radiotherapy and chemotherapy.
You can learn more about redox biology, free radicals and disease in the talk and video Harnessing free radicals: AI driven insights into lipid oxidation and new therapeutic strategies

The high sensitivity offered by BMG LABTECH readers like the PHERAstar, CLARIOstar Plus and VANTAstar for fluorescence measurements makes them powerful tools for monitoring cellular redox changes using biosensors like roGFP2. You can learn more about roGFP in the scientific talk and video Real-time monitoring of redox changes in cells with a microplate reader.

We have already highlighted some of the features of a microplate reader that aid in the study of metabolism in cancer cells in the applications just highlighted. Here we look closer at the technological benefits microplate readers bring to the table for the study of metabolism in cancer. Some of the essential instrument features relevant to the measurement of metabolism in cancer cell assays are summarized in Table 1.
Table 1. Summary of benefits offered by microplate readers for studies of metabolism in cancer cells.
| Feature | Benefit | Note |
| Ultrafast spectrometer | Captures full-absorbance data at a resolution of 1 nm for all wavelengths from 220 to 1000 nm in less than one second per well | Available on the PHERAstar FSX, VANTAstar and the CLARIOstar Plus, Omega series, and SPECTROstar Nano |
| LVF Monochromator | Provides wavelength flexibility with filter-like sensitivity for fluorescent and luminescent assays | Available on the VANTAstar and the CLARIOstar Plus |
| Enhanced Dynamic Range | Ensures accurate signal quantification across low to high concentrations of ligands and targets without running into the risk of signal saturation | Available on the PHERAstar FSX, VANTAstar and the CLARIOstar Plus |
| Atmospheric Control Unit | Independently control oxygen and carbon dioxide concentrations for cell-based assays (as low as 0.1% oxygen on the CLARIOstar Plus) | The VANTAstar, the CLARIOstar Plus, and the Omega series can be combined with the Atmospheric Control Unit for different kinds of live cell assays including cell viability, cell proliferation and other cell-based assays with specific requirements for the surrounding atmosphere. |
| Incubation and shaking | Optimize assay conditions with precise temperature incubation and shaking control, ensuring best results for cell-based and enzymatic applications | All BMG LABTECH readers offer accurate temperature regulation up to 45°C (some devices even offer the option of temperature regulation up to 65°C). Three shaking modes with adjustable speed up to 700 rpm (optionally to 1100 rpm) provide optimum aeration settings for cells that require agitation. The VANTAstar, CLARIOstar Plus, the Omega series and the SPECTROstar Nano can be equipped with an incredibly robust transport system for shaking 24/7 where required |
| Autofocus | By selecting autofocus at the start of a measurement, the reader automatically identifies the sample with the brightest light signal and automatically adjusts the focal height for this well | Available for BMG LABTECH microplate readers with Enhanced Dynamic Range function |
| Well scanning | Well scanning ensures uniform signal detection for cell-based and heterogeneous assays | Available on the PHERAstar FSX, VANTAstar and the CLARIOstar Plus, Omega series, and SPECTROstar Nano |
| Sampling rate | Sampling rate of 100 data points per second. Particularly useful for resolving fast kinetic processes and for generating higher density datasets of interaction events | Sampling rates for PHERAstar FSX and the CLARIOstar Plus |
| Reagent injectors | Reagent delivery to any plate format from 6-384 wells: control of injection timing, injection speed, delivery volume. Ability to inject different volumes in each well of a plate | The PHERAstar FSX, CLARIOstar Plus, and the Omega series allow simultaneous injection and measurement. The VANTAstar can be equipped with a modular injection unit. |
As we have seen, absorbance, luminescence and fluorescence measurements offer innovative ways to measure different assays for metabolism in cancer cells. Absorbance detection is available on BMG LABTECH’s complete portfolio of multi-mode microplate readers with the ultrafast spectrometer. BMG LABTECH’s multi-mode detection devices also include state-of-the-art capabilities for sensitive fluorescence and luminescence measurements. Multi-mode microplate readers bring benefits in speed, reproducibility and scale for routine and advanced measurements of metabolism in cancer cells.
Microplate readers offer quantitative and time-saving analysis for cell-based assays or in vitro biochemical-related assays. The use of different detection technologies means that researchers can deploy different techniques with varying sensitivities of measurement depending on the needs of their experimental systems.
Furthermore, the use of 96-, 384- or 1536-well microplates allows the processing of many samples simultaneously or in quick succession. This allows large numbers of samples to be measured in a single run which reduces the time for the collection of data. Real-time or time-resolved measurements are suitable to map the dynamic changes that may take place in metabolism in cancer cells. This applies to the kinetics of enzyme reactions and crucial measurements of the levels of metabolites.
Collectively, BMG LABTECH multi-mode readers combine high-quality measurements with miniaturized assays, short measurement times, and offer considerable savings on materials and other resources.
While the Warburg effect has been established for many years, our knowledge of the metabolism of cancer cells and how it may translate into new drugs and treatments is still a relatively new frontier. Therapies aimed at disrupting specific metabolic pathways in cancer cells can inhibit tumor growth and enhance the effectiveness of existing treatments. In the context of cancer therapy, targeting metabolic pathways has emerged as a promising strategy to exploit metabolic vulnerabilities in tumors. Indeed, several experimental drugs are under investigation that target key metabolic enzymes or pathways, such as glycolysis inhibitors and glutaminase inhibitors with some in early-stage clinical trials (as of writing April 2026). Among these, small molecule inhibitors targeting glycolytic enzymes and lactate transporters are showing potential to improve treatment efficacy and overcome chemoresistance.
Our understanding of metabolism in cancer cells continues to grow at pace, with trends in cancer research focusing on new metabolic pathways, oncometabolites, and therapeutic vulnerabilities. The demand for measurements related to changes in metabolism linked to cancer will therefore continue to increase in the years ahead as new discoveries, technologies and applications emerge from laboratories worldwide. Further advances in detection technologies and innovation in the specific assays available for microplate readers should serve as a priceless catalyst for further developments in studies of metabolism in cancer cells.
Wood WB et al. (1974) The molecular basis of metabolism. McGraw-Hill.
Atkinson DE (1977) Cellular energy metabolism and its regulation. Academic Press.
Stryer L (1975) Biochemistry. WH Freeman and Company, San Francisco.
Locasale JW, Cantley LC. Altered metabolism in cancer. BMC Biol. 2010 Jun 25;8:88. doi: 10.1186/1741-7007-8-88.
Vander Heiden MG, Cantley LC, Thompson CB. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science. 2009 May 22;324(5930):1029-33. doi: 10.1126/science.1160809.
Finley LWS. What is cancer metabolism? Cell. 2023 Apr 13;186(8):1670-1688. doi: 10.1016/j.cell.2023.01.038. Epub 2023 Feb 28.
Mukherjee S (2022) “The selfish cell” in The song of the cell. Simon & Schuster Inc. New York. pp.359-360.
Powerful and most sensitive HTS plate reader
Most flexible Plate Reader for Assay Development
Flexible microplate reader with simplified workflows
Upgradeable single and multi-mode microplate reader series
Absorbance plate reader with cuvette port











