PHERAstar FSX
Powerful and most sensitive HTS plate reader


Tobias Pusterla’s scientific background spans veterinary biotechnology, cancer cell biology, and the molecular mechanisms underlying inflammation‑driven tumorigenesis. After graduating in Veterinary Biotechnology at the University of Milan, Italy, he worked in mouse mutagenesis before completing a Ph.D. in Cellular and Molecular Biology through a joint program between the Open University of London, UK and the San Raffaele Scientific Institute, Milan, Italy. He later conducted postdoctoral research at the German Cancer Research Center (DKFZ) in Heidelberg, Germany, focusing on tumor biology, the tumor microenvironment, and the role of chronic inflammation in cancer development. His scientific work has contributed to understanding how damage‑associated molecular signals drive immune activation, cell migration, inflammation, and tumorigenesis, helping to clarify fundamental pathways linking cellular stress responses to physiological and pathological outcomes. After more than 13 years of research experience, he joined BMG LABTECH in 2013. Here, he oversees global marketing activities, including the creation of scientific content and the coordination of application support.
In the cell, several biological processes involve changes of the reduction-oxidation (redox) state, a misbalance of which can result in the production of free radicals and/or other Reactive Oxygen Species (ROS). Although, cells physiologically produce a certain amount of ROS (e.g.: respiratory chain in mitochondria), excessive production has the potential to significantly damage cell homeostasis, mainly through oxidative modifications of DNA/RNA and proteins. In several publications, this has been linked to diseases and aging.
Environmental conditions, such as exposure to heavy metals, can strongly influence oxidative stress. These environmental stressors modulate oxidative stress in different cellular compartments, making it important to measure redox potentials under various stress scenarios.
In order to protect themselves from non-physiological ROS concentrations, cells employ antioxidant molecules, mainly utilizing thiol-groups to catalytically detoxify themselves from free radicals. Generally, glutathione is the most abundant antioxidant molecule employed by the cell. It reduces ROS through the glutathione-peroxidase enzyme. The cytosolic glutathione buffer is normally highly reduced. Maintenance of a highly reduced glutathione redox potential is achieved through continuous reduction of glutathione disulfide by glutathione reductase (GR). The glutathione redox pair (GSH/GSSG) is crucial for redox homeostasis and redox signaling.
Redox processes are fundamental to the maintenance of cell homeostasis, influencing everything from energy production to cell signaling. At the heart of these processes is the glutathione redox potential, a critical indicator of cell redox homeostasis. Glutathione acts as a major antioxidant, helping to neutralize reactive oxygen species that are generated as natural byproducts of cellular metabolism. While ROS play important roles in redox signaling and defense mechanisms, their excessive accumulation can disrupt the delicate balance of redox potential within cells, leading to oxidative stress and cell damage. Monitoring these processes is essential for understanding how cells respond to environmental changes and stressors. The development of redox-sensitive probes, such as roGFP, has enabled researchers to track changes in glutathione redox potential and redox state in real time, providing valuable insights into the mechanisms that underpin redox homeostasis and the impact of oxidative stress on cell function.
The redox-sensitive GFP (roGFP) protein is a GFP variant that can be used as a biosensor for redox changes. In roGFP, two surface-exposed cysteines were introduced into the β-barrel (β-sheet reach region) of standard GFP at positions 147 and 204.
Importantly, roGFP has two different and redox-dependent excitation peaks at 405 nm and 488 nm, and one single emission peak at 510 nm. Measurement of excitation spectra at different redox states allows for characterization of the sensor’s redox-dependent changes. While cysteine oxidation results in an increase of the 405 nm excitation peak, cysteine reduction increases excitation at 488 nm.
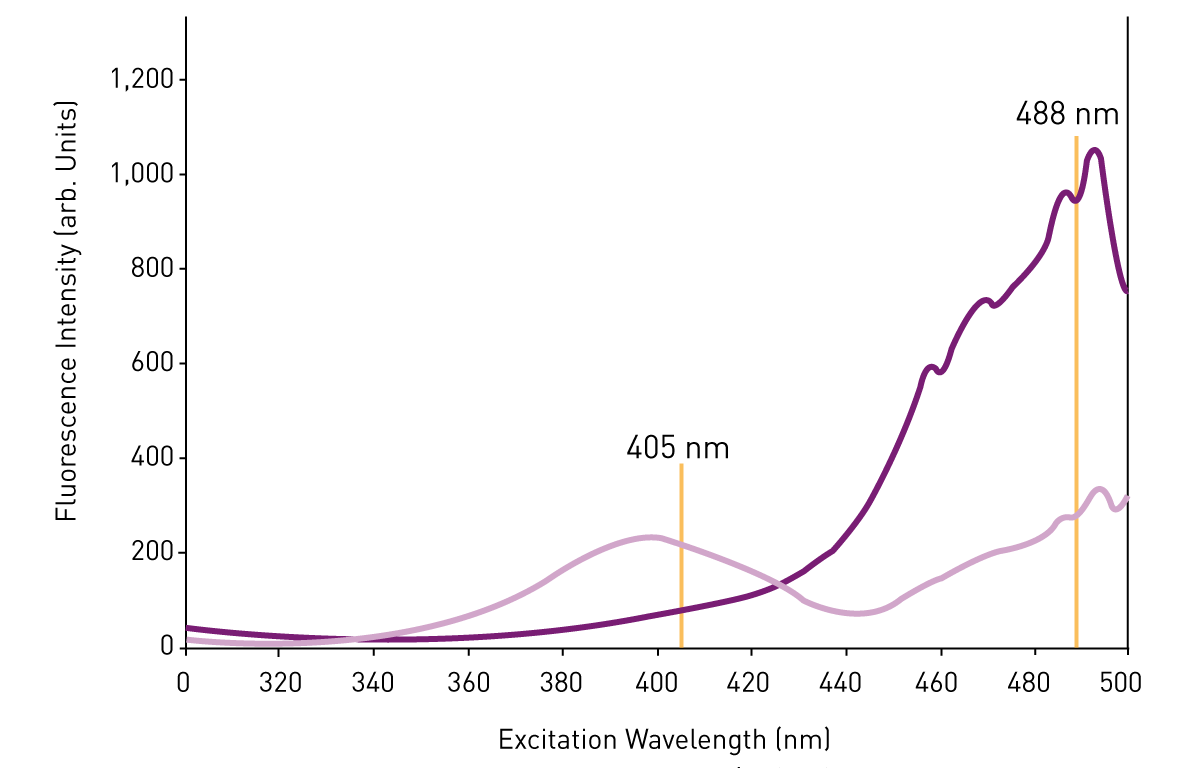
Consequently, the fluorescence ratio between emission at 510 nm and excitation either at 488 nm or 405 nm can be used as an indicator of the relative amount of oxidized/reduced roGFP. This fluorescence ratio reflects the redox state of the cell or cellular compartment in which roGFP is present, with the ratios for the two excitation peaks shifting in opposing directions depending on reduction or oxidation. Full oxidation of the sensor can be achieved by exposure to oxidizing agents such as H2O2, which is important for calibration and analysis of redox response.
roGFP is used to study the production of reactive oxygen species linked to cancer, neurodegenerative diseases, and aging. The roGFP1-R12 variant has been used to monitor redox potential in bacteria and yeast, as well as in multicellular organisms like C. elegans.
roGFP is genetically encoded and designed to detect subtle changes in the glutathione redox potential within cells. The unique mechanism of roGFP relies on the introduction of two exposed cysteines into the protein structure, which can form a reversible disulfide bond in response to the local redox environment. This structural change alters the fluorescence intensity of the protein, specifically shifting its two excitation maxima. roGFP can be targeted to specific cellular compartments, such as the mitochondrial matrix, allowing for detailed analysis of redox processes at the subcellular level.
The Grx1-roGFP2 fusion protein, which combines human glutaredoxin with roGFP2, further enhances sensitivity and dynamic range, making it particularly effective for monitoring rapid and subtle changes in glutathione redox potential across different cellular compartments. This versatility makes roGFP and its variants indispensable tools for studying redox processes and cell homeostasis in a wide variety of cell types.
Based on published data, roGFP has quite a few advantages:
There are several variants of roGFP that have been engineered to extend the range of glutathione redox potential measurements in vivo. Though, two versions are widely used: roGFP1 and roGFP2. Although both contain the same cysteine additions, roGFP1 is based on wild-type GFP whereas roGFP2 on eGFP. The main differences are that roGFP2 has a more efficient excitation at 488 nm than at 405 nm and offers a higher signal yield. On the other hand, roGFP1´s dynamic range extends more into the reduced range and is less pH-sensitive than roGFP2.
The specificity of roGFP2 for glutathione can be increased by linking it to human glutaredoxin 1 (Grx1), and the fusion of roGFP with human glutaredoxin 1 improves the kinetics of the redox reactions, allowing for faster response times in redox measurements.
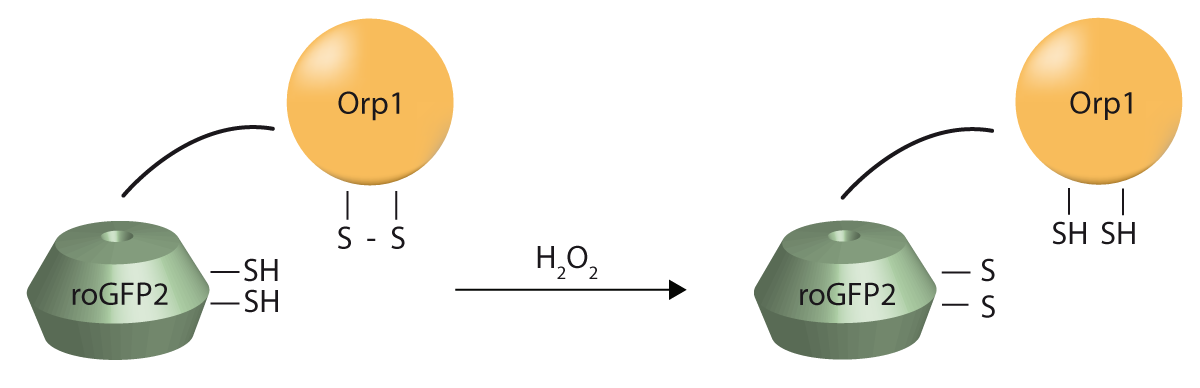
roGFP is usually expressed as fusion protein. Through this fusion, roGFP can be targeted to specific intracellular compartments and report for specific ROS in real-time. For instance, roGFP2-Orp1, the fusion of roGFP2 to the thiol peroxidase enzyme Orp1 is usually employed as a HO-sensitive probe and can be used to determine how chemicals induce or drugs decrease production of ROS in cells (AN252: Real-time monitoring of genetically encoded redox probes in mammalian cell monolayers).
In bacteria, roGFP is widely used to study redox potential and resistance mechanisms. In yeast, a specific roGFP biosensor targeting mitochondria can be used to compare ROS production in normoxic and hypoxic conditions (AN310: Mitochondrial oxidant generation follows oxygen deprivation and re-oxygenation).
Additionally, the expression of the glutathione-roGFP fusion complex (Grx1-roGFP) makes it possible to detect the glutathione-specific redox potential, allowing for real-time monitoring of fluctuations. For instance, the lab of Professor Bruce Morgan, Professor for Biochemistry at the University of Saarbrücken, Germany, uses this approach to understand the role glutathione plays in cell redox homeostasis and to better understand whether specific intracellular organelles preferentially store its oxidized or reduced form. Read here Prof. Morgan´s full interview.
In the scientific talk "Real-time monitoring of redox changes in cells with a microplate reader", Prof. Morgan discusses how redox-sensitive probes can be used to monitor redox enzyme activity. To address this question, roGFP2 was combined with glutaredoxin or glutathione. His team was then able to induce the oxidation of the signal molecule and to monitor in semi-high-throughput the change of roGFP fluorescence on a CLARIOstar Plus microplate reader.

Accurate measurement of redox processes using roGFP-based probes requires careful calibration and validation. Calibration is typically performed using well-defined redox buffers, such as dithiothreitol (DTT) for maximum reduction and hydrogen peroxide (H2O2) for maximum oxidation, to establish the probe’s response range under controlled redox conditions. This allows researchers to determine the probe’s dynamic range and calculate the degree of oxidation in living cells. Validation involves testing the probe’s response to various redox environments and comparing the results with established analytical methods, such as fluorescence detection in plate readers. Advanced imaging techniques, including confocal imaging, and high-throughput data acquisition with microplate readers, further support the validation process by enabling precise, real-time monitoring of roGFP fluorescence in different cell types and cellular compartments. These methods ensure that roGFP-based probes provide reliable and reproducible data on redox dynamics in cells.
Despite their transformative impact on redox biology, roGFP-based probes are not without limitations. Their fluorescence intensity can be influenced by environmental factors such as pH and temperature, which may affect the accuracy of redox measurements under certain conditions. Additionally, the presence of specific chemicals or drugs can interfere with probe function, necessitating careful experimental design and control. Calibration and validation are essential to account for these variables and to ensure that measurements reflect true redox processes rather than artifacts. Furthermore, the suitability of roGFP probes can vary between different cell types and subcellular compartments. Nevertheless, when properly applied, roGFP-based probes remain powerful tools for investigating redox signaling, oxidative stress, and cellular homeostasis in cells, offering unique insights into the dynamic nature of redox biology.
The best platform for ratiometric roGFP detection
BMG LABTECH´s microplate readers represent the ideal platform for the detection of roGFP-based biosensors in living cells. roGFP detection benefits considerably from sensitivity of detection in fluorescence intensity, measurements from the bottom of the plate, and from automated compound injection. “The CLARIOstar is sensitive enough that even without chemical perturbation, we are able to detect baseline sensor signals. For the redox community this is very, very important”, said Prince Saforo Amponsah about the role the CLARIOstar plays in his research. Watch below Prince´s full video interview.

In fact, the CLARIOstar and PHERAstar FSX gained a reputation as go-to plate readers in the roGFP community. The following features make them the ideal detection systems for roGFP-based assays:
Powerful and most sensitive HTS plate reader
Most flexible Plate Reader for Assay Development
Upgradeable single and multi-mode microplate reader series
Flexible microplate reader with simplified workflows
Binding constants quantify the strength of a binding reaction between a biomolecule and its target (ligand). But how do you measure them and what can you do with them?
Receptor-ligand kinetics is the study of the rates at which receptors and ligands interact, bind and dissociate. Learn why these types of measurements are important and how to measure them.
Receptor-ligand interactions are crucial for cell signalling. They are also important for drug discovery. How do microplate readers deliver benefits to both?