PHERAstar FSX
Powerful and most sensitive HTS plate reader


Barry Whyte is Application Scientist and Science Writer at BMG LABTECH in the United States. He has PhD and Bachelor of Science (BSc) degrees in biochemistry from the University of Bristol in the United Kingdom and more than 20 years of experience in the life sciences and science communications. Over the years, Barry has worked on three continents and traveled widely. He enjoys building on his international work experience and learning new ways to help scientists advance their research.
Prion diseases, Alzheimer’s, Parkinson’s, Huntington’s disease, and amyotrophic lateral sclerosis (ALS) are all part of the landscape of neurodegenerative diseases linked to protein misfolding. Each disease involves its own unique misfolded protein or combination of proteins leading to specific pathological features and clinical symptoms.
In this blog, we look at the relationship between protein misfolding and neurodegenerative disease. We also discuss how microplate readers can be used in neuroscience to help understand the impact protein misfolding has on disease.
Protein structure is closely tied to protein function.1 The structure of a protein is typically considered at four different levels: primary, secondary, tertiary, and quaternary. The primary structure is the sequence of amino acids in a polypeptide chain. Secondary structure refers to structures like alpha-helices and beta-sheets. Motifs of this type are local folded structures that form within a polypeptide chain due to interactions between atoms in the protein backbone.
Tertiary structure refers to the overall three-dimensional structure of a polypeptide. Many proteins are comprised of a single polypeptide chain and will only have primary, secondary and tertiary structures. However, some proteins like hemoglobin consist of several polypeptide chains that are arranged together to give the protein its quaternary structure.
Proteins are molecular machines. The sequence of amino acids in the three-dimensional structure of a protein determines its ability to bind to specific molecules, catalyze reactions, interact with other proteins, and carry out biological functions. Protein structure therefore impacts function at many different levels. This may include how a substrate binds to an enzyme’s active site or how effectively an enzyme catalyzes its reaction. It may also include the way proteins interact with other proteins or small molecules important for cell signaling.
Collectively protein structure has a huge impact on the overall stability and folding of a protein. It is therefore not surprising that changes in protein structure due to mutations, post-translational modifications or environmental factors can disrupt the functional properties of a protein and lead to altered or impaired function and disease.
The way proteins misfold and aggregate is closely associated with the accumulation of toxic proteins that cause many neurodegenerative diseases. Alzheimer’s involves the presence of two misfolded proteins in the brain: beta-amyloid protein and tau protein. Parkinson’s disease is typically characterized by the accumulation of the alpha-synuclein protein in the brain. Huntington’s disease is caused by an abnormal form of the huntingtin protein with an extended glutamine tract. Misfolded huntingtin protein forms amyloid aggregates that build up in neurons which in turn leads to neuronal dysfunction and cell death.
Prion diseases like Creutzfeldt-Jakob disease, bovine spongiform encephalopathy (BSE), scrapie, chronic wasting disease and kuru are caused by misfolded prion proteins. Prions have the innate ability to convert normal proteins into misfolded forms which makes prions infectious agents. Amyotrophic lateral sclerosis involves the misfolding and aggregation of several proteins including superoxide dismutase 1, TAR DNA-binding protein 43 (TDP-43) and fused in sarcoma (FUS). Aggregates of these proteins accumulate in motor neurons as hallmark cytoplasmic and nuclear inclusions. 2 
The neurodegenerative diseases mentioned earlier share a common theme: namely that the accumulation of misfolded proteins and the ensuing disruption of cellular processes contribute to the development and progression of disease. Many details remain to be worked out but the specific proteins involved in each disease adopt atypical conformations and fail to fold correctly. These misfolded proteins are often prone to aggregation and accumulate in the brain where they can have a myriad of effects including disruption of the function of organelles (for example mitochondria or the endoplasmic reticulum), impairment of protein degradation pathways, or alteration of the distribution of cellular components.
Misfolded proteins can also activate cellular stress responses, such as the unfolded protein response (UPR), oxidative stress pathways, and inflammatory responses. Stress responses are intended to protect cells and restore protein homeostasis. However, sustained, or excessive activation of these pathways can be detrimental to cellular health and contribute to disease.
Protein aggregates can also either directly or indirectly affect synaptic function. This type of interaction may interfere with the release and clearance of neurotransmitters, disrupt synaptic plasticity, impair the recycling of synaptic vesicles, and alter the composition of synaptic proteins. Disruptions of this type eventually contribute to the cognitive and motor deficits associated with disease.
Prolonged exposure to misfolded proteins and their amyloid aggregates induces cytotoxicity and triggers apoptopic or other cell death pathways in neurons. The outcome is a progressive, devastating loss of neurons and atrophy of the brain.
Protein misfolding can arise due to various factors including genetic mutations, environmental stress, post-translational modifications, chaperone dysfunction, imbalances in proteostasis, or conformational changes. This can lead to the cascading molecular events that impact health and which were described earlier. In some cases, mutations in the genes that code for proteins can directly affect their folding. These changes or mutations may alter the amino acid sequence or the structure of the protein making it more prone to misfolding. Examples include mutations in the presenilin genes in Alzheimer’s disease or mutations in the Huntingtin gene in Huntington’s disease.
The immediate environment is crucial to the integrity of proteins. Proteins are very sensitive to changes in their microenvironment and such alterations can lead to the exposure of hydrophobic regions of the protein, destabilization of protein structure, and subsequent misfolding. Changes in pH, temperature, or binding to other molecules may transition certain proteins from their native state to an alternative misfolded conformation.
Post-translational modifications, such as phosphorylation, glycosylation, or oxidation, can also impact protein folding. When these modifications go wrong, they can hinder the folding process and result in misfolded protein conformations.
Disruptions to protein homeostasis, which is also referred to as proteostasis imbalance, also generate detrimental effects on protein misfolding, degradation and clearance.
Chaperones are a group of functionally related proteins that assist the folding of proteins in the cell under physiological or stressed conditions. They help to prevent misfolding and promote the proper folding or refolding of partially unfolded proteins. When chaperones go awry, they too can lead to the formation of misfolded proteins. 3,4
Under normal conditions, chaperones interact with newly synthesized or partially folded proteins to provide a conducive environment for proper folding. They help prevent aggregation, promote correct folding pathways, and stabilize intermediate states of protein folding. When a protein fails to fold correctly, chaperones can bind to the misfolded regions and assist in the refolding process, helping the protein achieve its native structure. By keeping misfolded proteins in a soluble state, chaperones ensure proper folding or facilitate protein degradation through the quality control mechanisms of the cell.
In Alzheimer’s disease, protein misfolding is closely linked to the formation of beta-amyloid plaques. The initial step in the formation of these amyloid plaques involves the processing of the amyloid precursor protein. Amyloid precursor protein is a transmembrane protein normally cleaved by secretases to generate shorter, soluble fragments. In Alzheimer’s disease, a fraction of amyloid precursor protein undergoes aberrant processing through a series of enzymatic cleavages by beta-secretase and gamma-secretase. This abnormal processing leads to the production of longer forms of amyloid peptides, in particular beta-amyloid 42, which is more likely to misfold and aggregate. Misfolded beta-amyloid aggregates accumulate outside the neurons in the brain where they form amyloid plaques. The normal architecture of the brain is disrupted and neuroinflammation and neuronal damage are typical.
The accumulation of amyloid plaques contributes to neurotoxicity and promotes the development of other pathological features of Alzheimer’s disease such as neurofibrillary tangles composed of hyperphosphorylated tau protein. The interplay of amyloid plaques, tau pathology, neuroinflammation, and other cellular processes ultimately leads to synaptic dysfunction, neuronal loss, and cognitive decline.
In Parkinson’s disease, protein misfolding is associated with the onset and progression of the disease as well as the formation of Lewy bodies. The misfolding of alpha-synuclein is a key event in this process. In Parkinson’s disease, alpha-synuclein adopts an abnormal conformation and subsequently misfolds, which leads to the formation of small soluble aggregates or oligomers. Further aggregation leads to the formation of larger insoluble aggregates or fibrils that are typical of Lewy bodies. The net result is disruption of several cellular processes including regular protein degradation, mitochondrial function, synaptic activity, and intracellular trafficking. Cellular dysfunction and neuronal damage ensue.
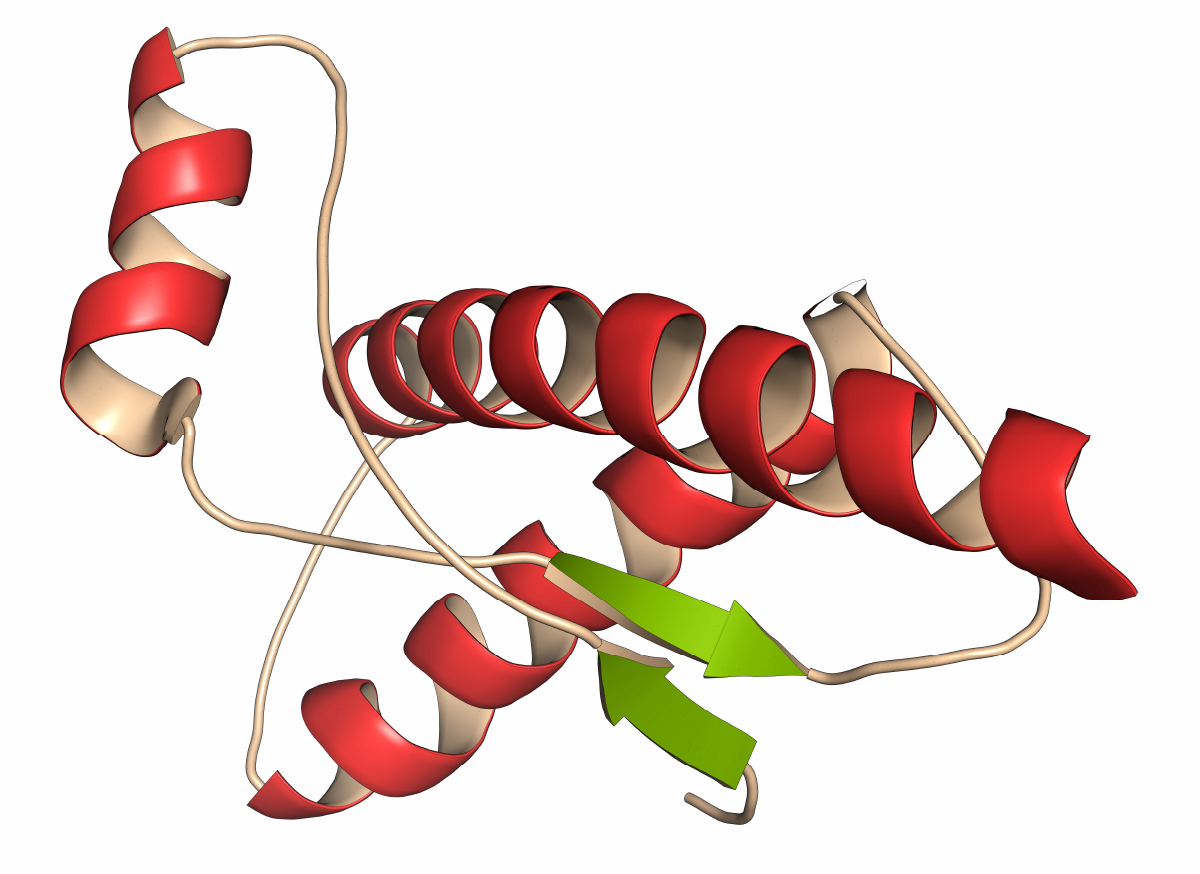 Protein folding plays a critical role in prion diseases including chronic wasting disease (CWD), scrapie, bovine spongiform encephalopathy (BSE), Creutzfeld Jakob Disease (CJD), and Kuru.
Protein folding plays a critical role in prion diseases including chronic wasting disease (CWD), scrapie, bovine spongiform encephalopathy (BSE), Creutzfeld Jakob Disease (CJD), and Kuru.
Prion diseases are caused by the conversion of the naturally occurring PrPC prion protein into the misfolded form PrPSc. PrPSc has a different conformation to PrPC and contains a high content of beta-sheets. The infectious form of prions can be transmitted by ingestion or other types of exposure to the protein. Once transmitted, PrPSc will continue to propagate and lead to the generation of abnormal conformations in other prion proteins. One of the consequences of PrPC transmission is protein aggregation and amyloid accumulation which in turn leads to neurotoxicity. Changes take place in a few months to several years and are ultimately fatal. The brains of infected organisms typically contain large vacuoles making them spongy in nature.
Huntington’s disease arises from a mutation in the huntingtin gene (HTT) that generates an expanded CAG repeat in its protein-coding region. Huntingtin proteins thus have an extended tract of glutamine amino acids that may contain from around 36 to 250 additional glutamine residues. This extended tract leads to conformational changes that produce misfolding of the protein with dramatic consequences for cellular function. Huntington’s disease typically leads to progressive changes in the functional abilities of patients and usually results in movement, cognitive and psychiatric disorders. These changes arise due to a progressive breakdown of nerve cells in the brain.
Protein misfolding and subsequent aggregation are also implicated in amyotrophic lateral sclerosis, a potentially fatal disease that affects nerve cells in the brain and spinal cord.
ALS, which is also known as Lou Gehrig’s disease, worsens over time and leads to loss of muscle control. Several proteins are linked to ALS including superoxide dismutase 1 (SOD1), valosin-containing protein, ubiquilin-2, p-62, Transactive Response (TAR) DNA-binding protein 43 (otherwise known as TDP-43) and fused in sarcoma/translocated in liposarcoma (FUS/TLS). Misfolded versions of these proteins have a high likelihood to aggregate which disrupts cellular function, leads to toxicity and degeneration of motor neurons, and can spread to proximal regions in the brain.
Microplate readers are used widely to study protein misfolding. Enzyme-linked immunosorbent (ELISA) assays, cell-based assays, fluorescence-based applications, protein stability assays and protein-protein interaction studies are some of the options researchers turn to when they want to investigate the way proteins misfold. ELISA assays are used to detect and quantify misfolded protein species or specific markers linked to protein misfolding. The presence of misfolded proteins can be measured in samples using specific antibodies that specifically recognize misfolded conformations or post-translational modifications.
In the study “Seeded propagation of α-synuclein aggregation in mouse brain using protein misfolding cyclic amplification” a team of researchers used protein misfolding cyclic amplification (PMCA) to study the aggregation of alpha-synuclein in mouse brain tissue.5 ELISA assays were performed on a CLARIOstar® Plus microplate reader to detect Ser129 phosphorylated alpha-synuclein. PMCA faithfully replicates mammalian prions in vitro. After seeding with a small quantity of a sample containing the pathologic prion protein, the method allows conversion of the cellular prion protein from a normal brain substrate into an aggregated form using repeated cycles of sonication and incubation. The authors demonstrated the in vitro generation and seeded propagation of alpha-synuclein aggregates in mouse brain tissue and subsequent transmission of an experimental synucleinopathy in M83 mice by intracerebral inoculation.
Microplate readers equipped with fluorescence detection capabilities are frequently used to probe protein misfolding. Fluorescent dyes, such as thioflavin T or Congo red, bind to misfolded amyloid aggregates and result in an increase in fluorescence signal. By monitoring the fluorescence intensity or changes in fluorescence over time, researchers can assess the kinetics of protein aggregation and evaluate the effects of different conditions or compounds on protein misfolding.
A paper entitled “RT-QuIC detection of CWD prion seeding activity in white-tailed deer muscle tissues” demonstrates how RT-QuIC assays can be performed on a FLUOstar® Omega microplate reader to characterize prion-seeding activity in samples of tissue from wild and farmed deer.6 The ThT fluorescence-based assays successfully detected prions responsible for chronic wasting disease in the muscles of infected animals. The authors report that the RT-QuIC assay described in the study could be a useful tool to monitor CWD prions in venison and could be used to reduce and/or prevent CWD prions from entering animal and human food chains.
In the paper “Assessment of Real-Time Quaking-Induced Conversion (RT-QuIC) Assay, Immunohistochemistry and ELISA for Detection of Chronic Wasting Disease under Field Conditions in White-Tailed Deer: A Bayesian Approach”, a team of scientists looked at the relative merits of using different assays to detect chronic wasting disease in infected animals.7 The authors compared a fluorescence-based ThT assay with gold standard methods of immunohistochemistry and ELISA. The RT-QuIC assays were performed on a FLUOstar® Omega microplate reader. Immunoassays are highly effective in detecting infected animals with chronic wasting disease, particularly those in the later stages of disease. The RT-QuIC assays described in the study offered high sensitivity and specificity for detecting the presence of infective prions in biological samples. Moreover, the RT-QuIC assay was well suited for tests in early-stage infected animals and in conditions of natural transmission.
You can learn more about RT-QuIC assays, including how to optimize the sensitivity and specificity of RT-QuIC measurements, in the following video:

Fluorescently tagged misfolded proteins can also be expressed in cells which makes them amenable to cell-based assays on microplates. The aggregation of labeled proteins can be quantified using fluorescent intensity measurements. These types of assays provide insight into the cellular processes involved in protein misfolding and can also be used to screen for drugs for disease interventions.
 In the paper “Time-resolved FRET screening identifies small molecular modifiers of mutant Huntingtin conformational inflexibility in patient-derived cells”, researchers demonstrated that a Time-Resolved FRET assay could be miniaturized and applied to a compound screening workflow for cells of patients with Huntington’s disease.8 The signals from the well-established antibody-based assays were detected on a PHERAstar® FSX microplate reader. After cell lysis, the assay quantifies conformational flexibility changes for mutant Huntingtin protein by measuring the spatial proximity of an N-terminal region of the protein relative to specific antibodies. The method is suited to large high-throughput screening campaigns to identify previously unknown Huntington’s disease drugs and drug targets.
In the paper “Time-resolved FRET screening identifies small molecular modifiers of mutant Huntingtin conformational inflexibility in patient-derived cells”, researchers demonstrated that a Time-Resolved FRET assay could be miniaturized and applied to a compound screening workflow for cells of patients with Huntington’s disease.8 The signals from the well-established antibody-based assays were detected on a PHERAstar® FSX microplate reader. After cell lysis, the assay quantifies conformational flexibility changes for mutant Huntingtin protein by measuring the spatial proximity of an N-terminal region of the protein relative to specific antibodies. The method is suited to large high-throughput screening campaigns to identify previously unknown Huntington’s disease drugs and drug targets.
Scientists have faced many challenges in the development of drugs targeting protein misfolding due to uncertainty about which protein is responsible for the different neurodegenerative diseases. Researchers also have faced many hurdles in translating preclinical findings to clinical trials for diseases associated with misfolded proteins. 3
Attempts have focused on targeting the production, misfolding and aggregation of key proteins. The application note ‘Peptidic inhibitors of α-Synuclein that prevent Parkinson-associated fibrilization and cytotoxicity’ highlights one such approach with the testing of peptide-based inhibitors that decrease the rate of nucleation and fibril elongation. Efforts have also been directed at targeting chaperones, degradation processes, extracellular clearance, and rebalancing the proteostatic network.
Chaperones, which are involved in all aspects of proteostasis, make a good entry point for interventions. Small molecule inhibitors or activators have been sought for different disease targets. A second approach has been to protein engineer chaperones to improve their ability to breakdown aggregated proteins.
The ubiquitin proteasome system is an example of one of the protein degradation processes where interventions have also been sought. Several small molecules have been identified that promote the degradation of disease proteins in vitro but in most cases these efforts and molecules have not translated well to in vivo tests. Inhibition of the mammalian target of rapamycin (mTOR) has been a popular target in drug discovery activities and is effective at stimulating the degradation of neurodegenerative disease proteins. However, mTOR regulates many other processes beyond protein degradation which makes it challenging to come up with a drug of the required specificity.
Many attempts at targeted protein clearance have focused on the use of antibodies to promote clearance of beta-amyloid. Uncertainty about which antibodies engage clinically relevant forms of amyloid remains although there has been some recent progress in direct targeting by monoclonal antibodies (aducanumab and donanemab).
Rebalancing the proteostatic network is another therapeutic strategy being pursued. Overexpression of human heat shock factor 1 is protective of neurons in cell models of neurodegenerative disease. It reduces polyglutamine aggregate formation and prolongs lifespan in a mouse model of Huntington’s disease. However, the effects of heat shock factor drugs are likely to have other effects including tumorigenesis under some conditions. The hope is to harness the neuroprotective effects independent of the tumorigenic effects, but further work is needed to make this a reality. 9
New drugs are actively being sought for the different targets highlighted in this blog. In clinical trials, there is a big need for translatable biomarkers that track the progression and severity of the neurodegenerative diseases impacted by protein misfolding. Some clinical trials have recently made progress for targeting beta-amyloid in Alzheimer’s disease, but much work lies ahead. Progress in artificial intelligence and modeling techniques have made significant contributions to the study of protein folding and misfolding which should bring further advances to the drug discovery space.
The PHERAstar FSX was specifically conceived for screening campaigns and is your go-to reader for high-performance high-throughput screening.
Both the VANTAstar® and CLARIOstar Plus allow for wavelength flexibility and include Enhanced Dynamic Range technology for superior performance in a single run. They also offer increased light transmission and sensitivity courtesy of Linear Variable Filter MonochromatorsTM and different filter options.
All BMG LABTECH microplate readers have exceptionally fast reading capabilities. In addition, the Omega series, CLARIOstar Plus and PHERAstar FSX microplate readers come with on-board injectors that can offer the very best options for detection at the time of injection.
BMG LABTECH’s Omega series of readers are a preferred choice for RT-QuIC aggregation assays as they have the robustness to withstand extensive and prolonged shaking.10
Collectively, these multi-mode readers combine high performance with miniaturized assays, short measurement times, and offer considerable savings on materials and other resources.
Configure your microplate reader and get an initial recommendation!
Powerful and most sensitive HTS plate reader
Most flexible Plate Reader for Assay Development
Upgradeable single and multi-mode microplate reader series
Flexible microplate reader with simplified workflows
Neurodegenerative disease ultimately leads to the death of neurons as neuronal functions deteriorate. Find out how microplate readers can be used to study neuronal cell death and its link to neurodegenerative disease.
The disruption of mitochondrial function is linked to neurodegenerative diseases. Find out how microplate readers advance research into mitochondrial dysfunction and neurodegenerative diseases.
Alpha-synuclein is a key protein involved in neurodegenerative diseases like Parkinson’s. Find out how microplate readers can help advance alpha-synuclein research.
Amyloids are thought to play a crucial role in neurodegeneration. Find out how microplate readers help advance amyloid research.
The tau protein plays a role in many neurological diseases and disorders. Find out about neuronal toxicity induced by tau and how microplate readers can aid tau research.