Introduction
To survive as sessile organisms, plants need to constantly adapt their metabolism to their environment. Flooding of plants is widespread and has severe metabolic consequences as it limits the cellular supply with oxygen to drive respiration, and thus production of ATP. Submergence acclimation typically involves drastic alterations in metabolism to circumvent anoxia and to maintain primary metabolism and ATP supply. We have recently established the use of a genetically encoded protein sensor (ATeam1.03-nD/nA, see Ref. 1) to assess MgATP dynamics in the model plant Arabidopsis thaliana. This system allows for reliable analyses of changes in MgATP in vivo and in real-time and provides a new handle to investigate the energetic consequences of low oxygen for the plant cell.
Assay Principle
ATeam is a genetically encoded Förster resonance energy transfer (FRET)-based protein sensor that reversibly binds MgATP1. MgATP binding induces a structural change of the MgATP-binding domain and results in more efficient FRET from the blue fluorophore mseCFP (CFP) to the yellow fluorophore cp173-mVenus (Venus). (Figure 1A). High Venus/CFP fluorescence emission ratios thus indicate high MgATP levels and vice versa (Figure 1B). ATeam can be expressed stably, consitutively and at high levels in Arabidopsis plants, and low oxygen stress can be imposed by submerging plants while simultaneously restricting their oxygen supply (Figure 1C).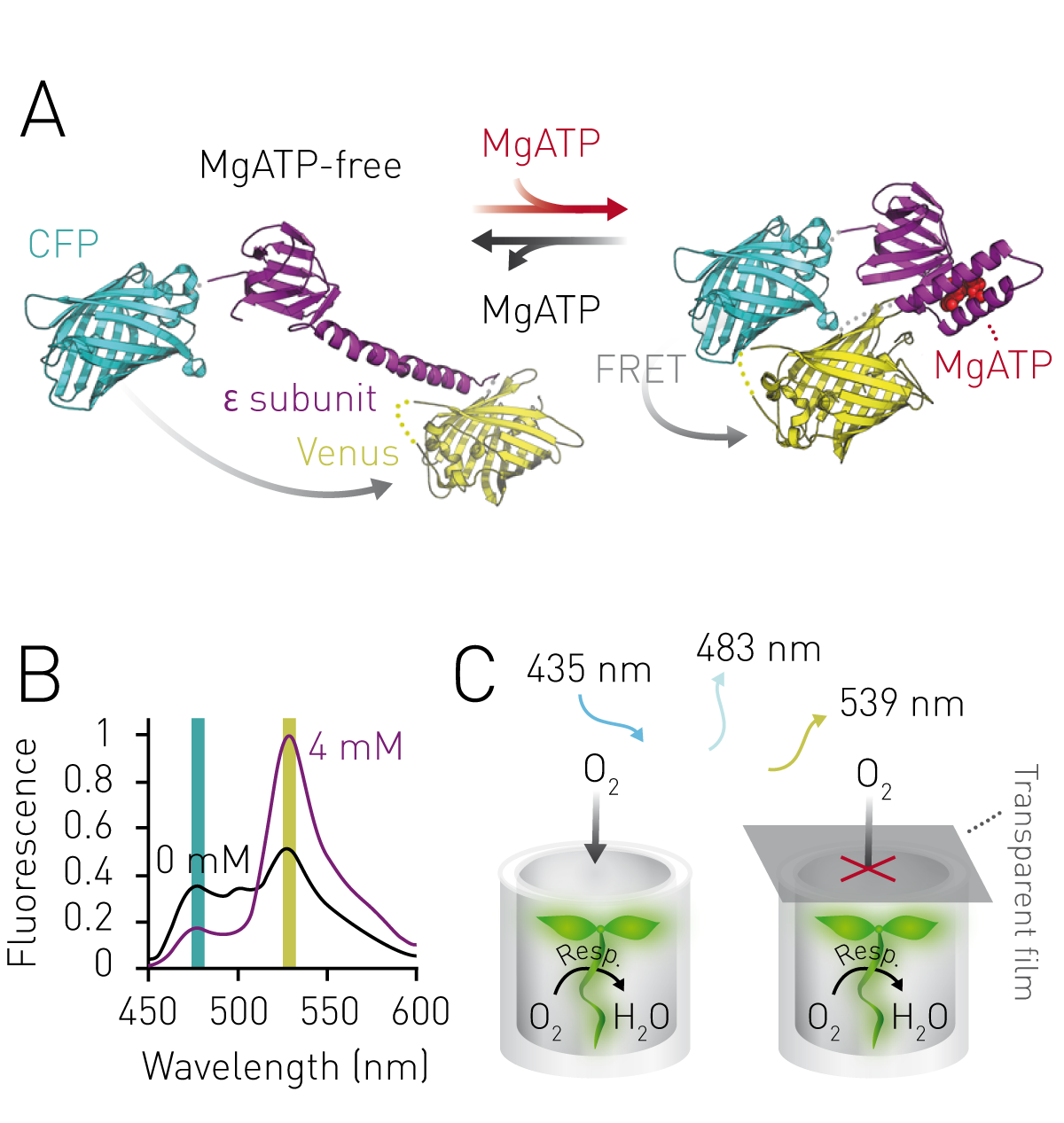
Materials & Methods
- CLARIOstar microplate reader (BMG LABTECH)
- Transparent Microtest Plate 96 Well,F (Sarstedt #82.1581)
- Transparent film for qPCR (VWR #391-1258)
- Assay medium (10 mM MES, pH 5.8, 10 mM CaCl2, 10 mM MgCl2, 5 mM KCl)
Experimental procedure
Arabidopsis seedlings were submerged in assay medium on a transparent 96 well microtest plate. Wells were either left unsealed (control condition allowing for resupply of the medium with atmospheric oxygen) or sealed with a transparent film to restrict resupply of the assay medium with oxygen. The ATeam fluorescence was recorded while - except from the excitation flashes - the plate was kept in the dark to suppress oxygen evolution through photosynthesis.
Instrument settings
| Optic settings | Fluorescence intensity, plate mode kinetic, top optic | |
| Number of multichromatics 2 | ||
|
Monochromator settings
|
CFP (gain 1250)
Excitation 435-20 nm Dichroic 459.5 Emission 483-18 Venus (gain 1200) Excitation 435-20 nm Dichroic 488.8 Emission 539-13 |
|
| Well scan | Orbital averaging with a 3 mm diameter | |
| General settings | Number of flashes per cycle 50 | |
|
Settling time 0.2 s
|
||
| Kinetic settings | Number of cycles 800 | |
| Cycle time 180 s | ||
| Shaking | 10 s double orbital shaking at 400 rpm before each cycle | |
Results & Discussion
Arabidopsis seedlings exposed to low oxygen stress showed gradually decreasing Venus/CFP ratios in the cytosol (Figure 2A), while the values remained almost constant in seedlings that were submerged in assay medium but not sealed to allow for oxygen resupply, except for an initial decrease immediately following seedling immersion (Figure 2B). This suggests that MgATP supply to the cytosol under oxygen-limiting conditions is not able to balance cytosolic ATP consumption. Unsealing of wells after 16 h led to a recovery of MgATP levels through re-oxygenation of seedlings, demonstrating the reversibility of ATeam readings (Figure 2C). Individual emission intensity plots of Venus and CFP highlight a key advantage of ratiometric FRET-based protein sensors over intensiometric sensors and/or dyes: while in Figure 2B individual fluorophore intensities vary (e.g., through sample movement), the Venus/CFP ratio remains relatively stable. A true difference in MgATP binding as seen in Figure 2A is, in contrast, characterized through the antiparallel behaviour of both fluorophore emission intensities as a result of bona fide FRET.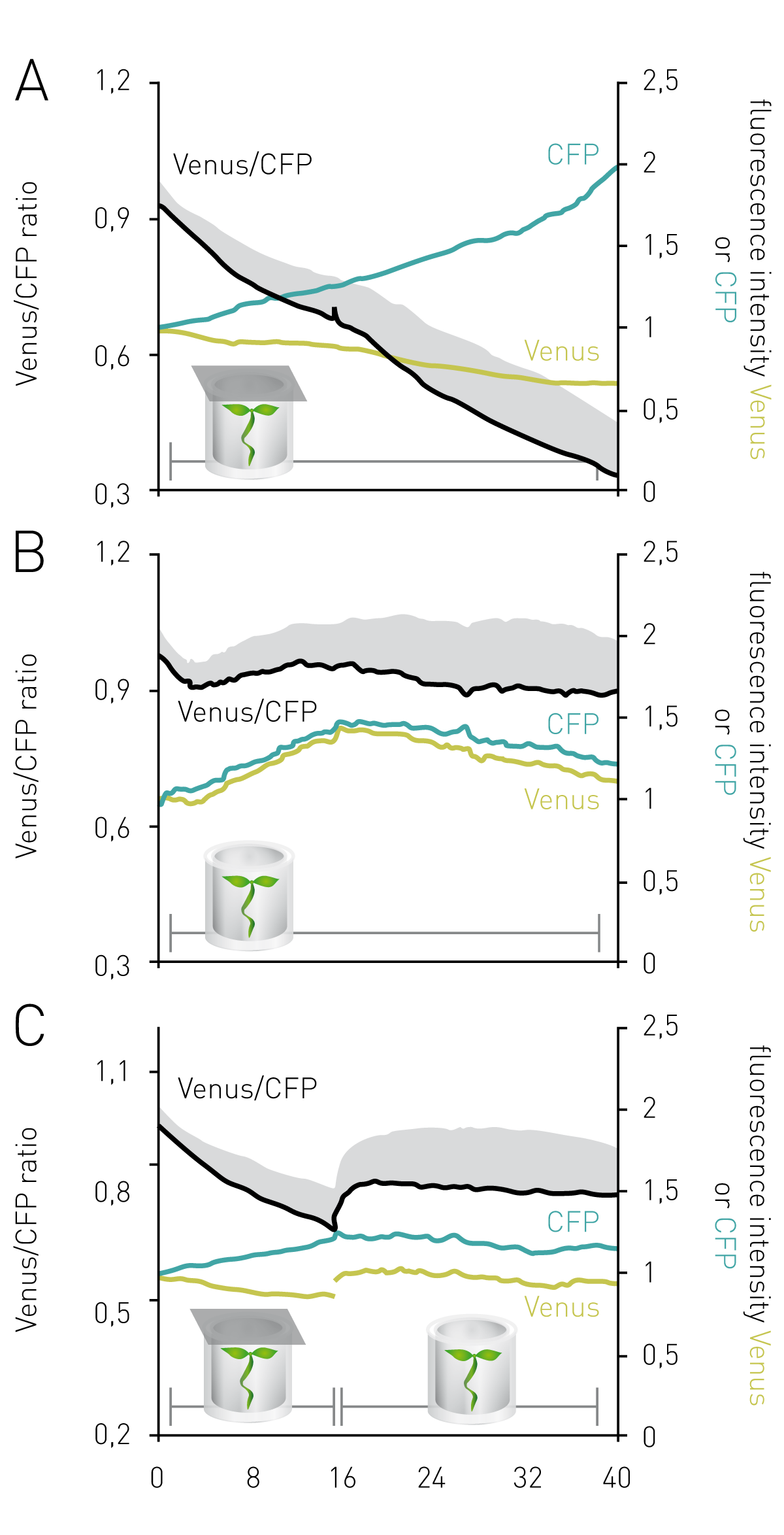
Conclusion
The CLARIOstar microplate reader allows to read the fluorescence of genetically encoded protein sensors (e.g., ATeam1.03-nD/nA) expressed in plants exposed to naturally occurring stress conditions (e.g., hypoxia as a hallmark of submergence stress).
References
- Imamura, H et al. (2009) Visualization of ATP levels inside single living cells with fluorescence resonance energy transfer-based genetically encoded indicators. Proc. Natl. Acad. Sci. USA 106: 15651-15656.
- De Col, V et al. (2017) ATP sensing in living plant cells reveals tissue gradients and stress dynamics of energy physiology. eLife 6: e26770.