Introduction
Salmonella enterica serovar Typhimurium are amongst the leading causes of gastrointestinal disease. Multiple antibiotic resistance (MAR) in salmonella has been increasing and over 30% of isolates of S. Typhimurium. Efflux is one mechanism that can confer MAR, efflux pumps are membrane proteins that actively export a wide range of toxic substrates including antibiotics, dyes, and biocides to the cells’ external environment, thereby preventing accumulation of toxic agents and mediating resistance to these agents as a result.
The wide spectrum of substrates recognized by efflux systems has prompted concern that exposure of a bacterium to one substrate could select for over-expression of an efflux system and consequent resistance to all other substrates. It is possible that exposure of a bacterium to a biocide could result in selection of an efflux mutant which has reduced susceptibility to antibiotics, which it has not previously encountered. In this study, mutants of salmonella resistant to biocides were selected. The growth rates of biocide resistant mutants were determined and compared to parent strains and the effect of the addition of biocides during growth was determined.
Materials & Methods
Selection of biocide resistant mutants
Previously, a panel of Salmonella Typhimurium was used to select biocide resistant mutants. SL1344 was used as a reference control strain, L108 is a derivative of SL1344 lacking the TolC porin and L358 is a multiple antibiotic resistant DT104 isolate. L378 is a cipro-floxacin resistant isolate from poultry and L357 is a representative DT104 isolate. The biocides AQAS (a quaternary ammonium compound), Superkill (a mix of aldehydes), Farm Fluid S (a mixture of tar oils and phenolics), and Virkon S (peroxygen generating compound) were used to select for tolerant mutants by incorporation in agar and overnight incubation after inoculation with each strain.
Analysis of growth kinetics of biocide tolerant mutants
The rate of growth was determined over 24 h at 37°C using a BMG LABTECH microplate reader. 100 μL of sterile LB broth was dispensed into clear sterile 96-well microplates and inoculated with overnight culture of each strain to give a final inoculum of 4%. Readings were taken every ten minutes of absorbance of each well (scanned at 600 nm) in the microplates over the 24 h time period. Each strain was analysed in triplicate wells on at least three separate occasions to give nine data sets for analysis. Additionally, each strain was challenged with either 0.5 X, or the MIC (minimum inhibitory concentration) of the selective biocide for the parent after two hours growth in order to determine whether the inhibitory ability of each biocide was reduced in mutants, respective to parent strains.
Results & Discussion
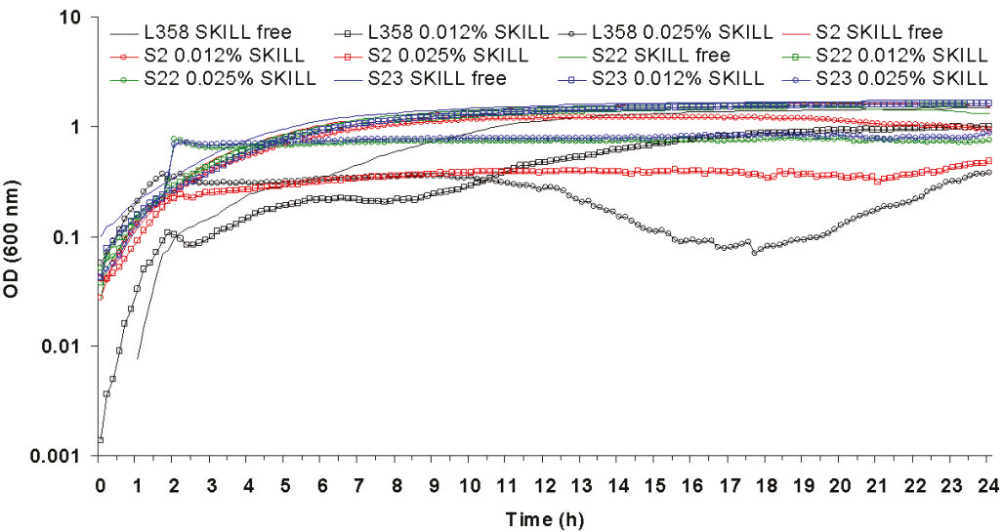
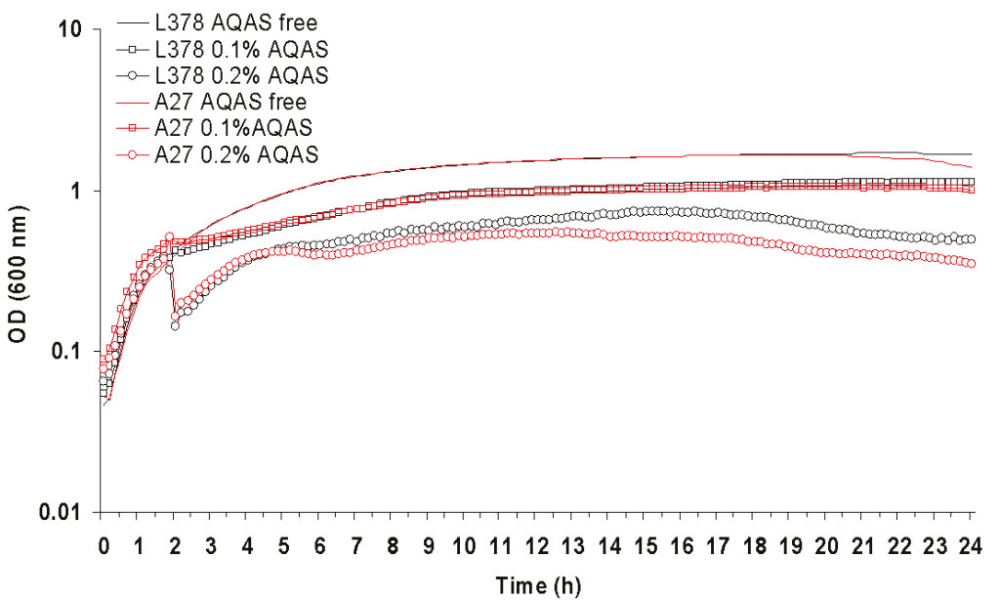
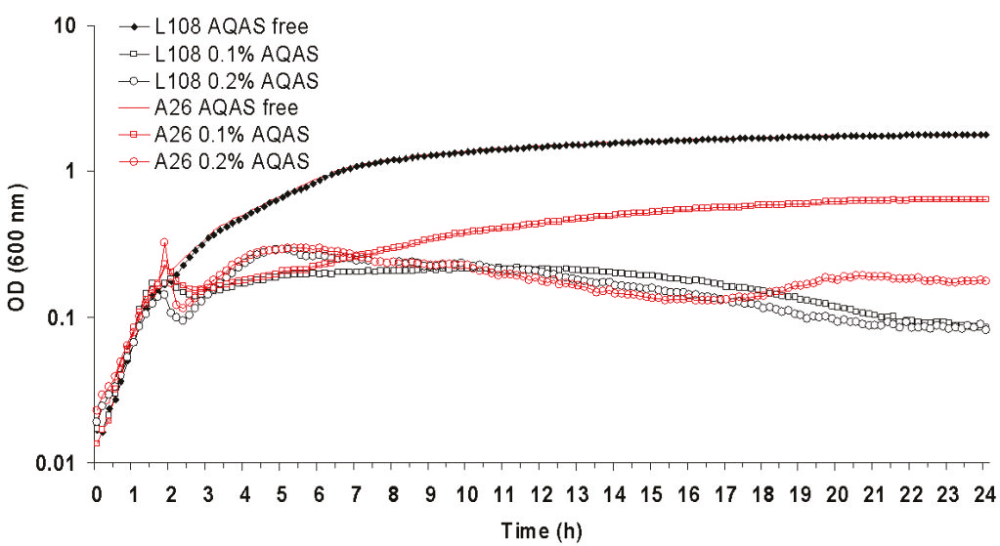
The majority of biocide selected mutants were more resistant to the addition of biocides to the media than their respective parent strains, including those strains for which the MIC of the selective biocide had remained unchanged when compared to the parent (table 1). AQAS selected mutant A27 and A26 were able to grow significantly (p >0.01) better upon both the addition of 0.5 X and the MIC of L378 and L108, respectively, to the media (figures 2 and 3).
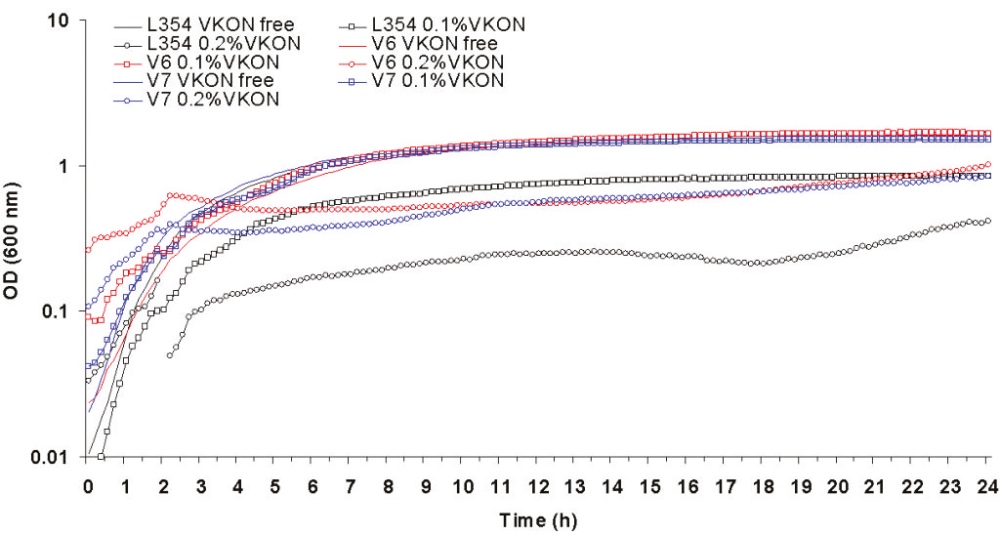
Virkon selected mutants V6 and V7 both grew significantly (p > 0.01) better than L354 when challenged with 0.5 X or the MIC of Virkon for L354 (figure 4).
 No significant differences were observed between Farm Fluid S selected mutants F1 and F2 and their parent L357 in biocide free broth or when exposed to 0.5 X the MIC of Farm Fluid S. However both mutants grew significantly (p > 0.05) better when challenged with the MIC of Farm Fluid S (0.1% and 0.2% final concentration) as shown in figure 5.
No significant differences were observed between Farm Fluid S selected mutants F1 and F2 and their parent L357 in biocide free broth or when exposed to 0.5 X the MIC of Farm Fluid S. However both mutants grew significantly (p > 0.05) better when challenged with the MIC of Farm Fluid S (0.1% and 0.2% final concentration) as shown in figure 5.
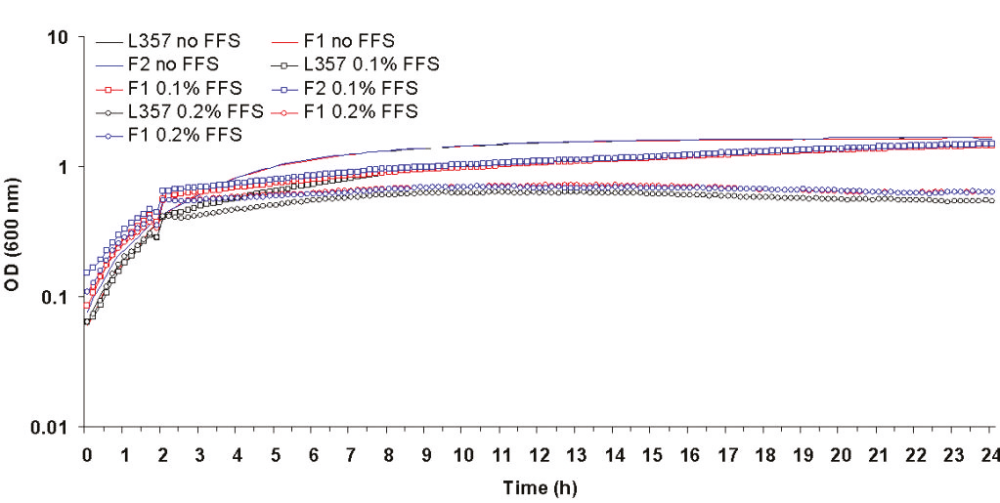 Table 1: Parent strains are in bold and mutants are listed below their respective parents. Mutants with increased resistance to biocide compared to their parents are indicated in red.
Table 1: Parent strains are in bold and mutants are listed below their respective parents. Mutants with increased resistance to biocide compared to their parents are indicated in red.
| MIC (%) | |||||
| Strain and Genotype | Selective agent |
Virkon |
Superkill | AQAS | FFS |
| L357 (DT104 ‘A’) | 0.4 | 0.025 | 0.1 | 0.2 | |
| F1 | FFS (1X MIC) | 0.2 | |||
| F2 | FFS (1X MIC) | 0.2 | |||
| L378 (CipR VLA52) | 0.4 | 0.025 | 1.6 | 0.2 | |
| A27 | >3.2 | ||||
| L108 (tolC::aph from SL1344) | 0.4 | 0.006 | <0.003 | 0.025 | |
| A26 | AQAS (2X MIC) | 0.12 | |||
| L358 (MDR DT104) | 0.4 | 0.025 | 1.6 | 0.2 | |
| S2 | S’kill (1X MIC) | 0.025 | |||
| S22 | S’kill (2X MIC) | 0.025 | |||
| S23 | S’kill (2X MIC) | 0.025 | |||
| SL1344 | 0.4 | 0.025 | 0.1 | 0.2 | |
| V6 | V’kon (2X MIC) | 0.8 | |||
| V7 | V’kon (2X MIC) | 0.8 | |||
Conclusion
This data clearly indicates that biocide exposure selects for strains with increased tolerance to biocides at sub-MIC concentrations and at the MIC and that there is no obvious fitness cost in these strains when compared to their parents in biocide free broth.