Introduction
Highly ordered protein aggregates, termed amyloid fibrils, are associated with a broad range of diseases, many of which are neurodegenerative, for example, Alzheimer’s and Parkinson’s. The transition from soluble, functional protein to insoluble amyloid fibril occurs via a complex process involving the initial generation of highly dynamic early stage aggregates or prefibrillar species. Prefibrillar species (dimers, tetramers etc.) are proposed to play a key role in the cytotoxicity of amyloid fibrils. Therefore, novel probes that have broad applicability in the detection of prefibrillar species of amyloidogenic proteins are actively being sought.
We have recently reported1 a novel broad-spectrum fluorescent probe: (bis(triphenylphosphonium) tetraphenylethene (TPE-TPP)), with emission characteris tics that are eminently suited to monitoring prefibrillar aggregation of various protein species using various fluorescence techniques. Herein, we describe our results for Fluorescence Polarisation (FP).
Assay Principle
FP measures the rotation of molecules in solution, using plane polarised light. Generally, small fluorophores such as TPE-TPP rotate quickly in water. When excited with polarised light, the individual molecules rotate randomly before emission. Their emitted light is depolarised and has a small FP value. If the fluorophore is bound to a large protein the whole complex rotates slowly. The emitted light from the complex remains plane polarised and the FP value is large (Fig. 1). 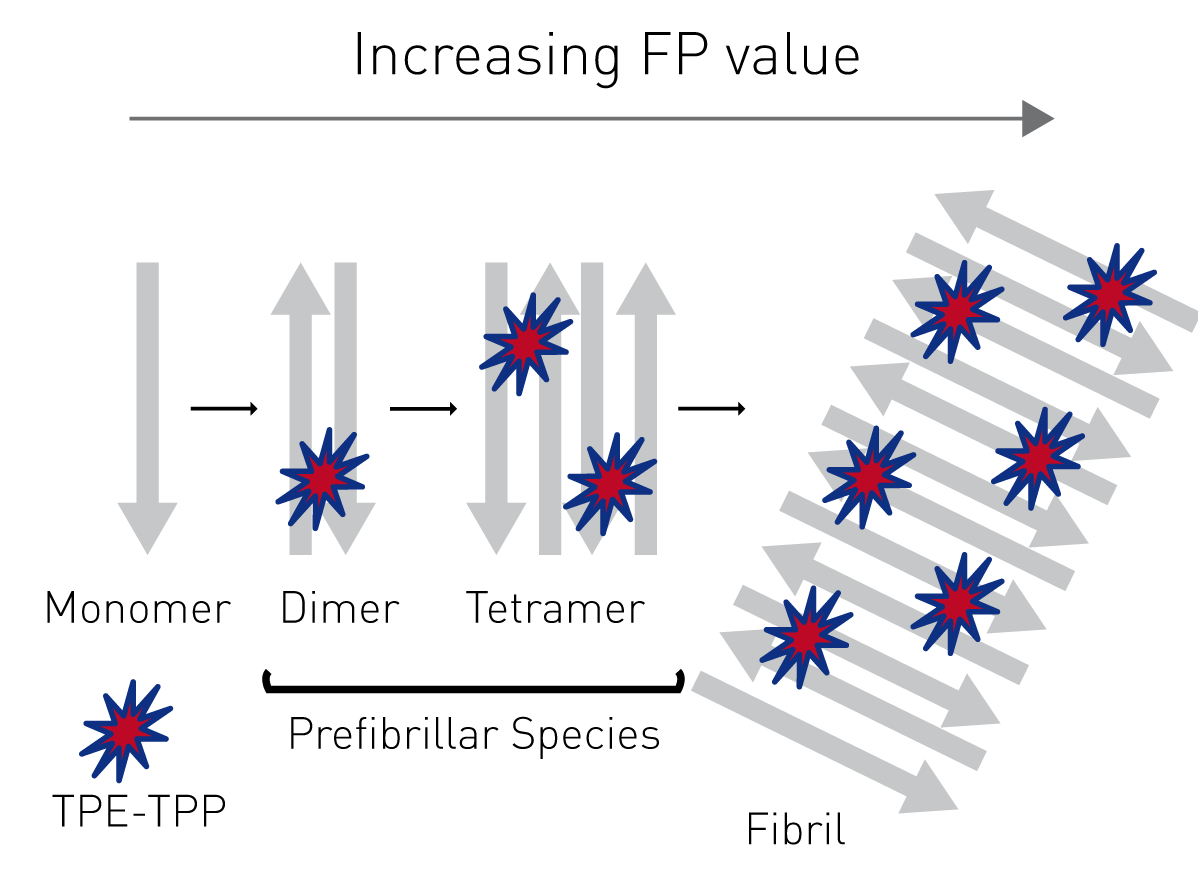
This technique is well suited to study amyloid aggregation of proteins as the FP value is dependent on the increasing mass of the TPE-TPP/protein complex and less dependent on the fluctuation of fluorescence intensity caused by the inhomogeneity of the solution. In this study, the change in FP for TPE-TPP during the aggregation of reduced and carboxymethylated α-lactalbumin (RCM α-LA), RCM ĸ-casein and insulin, was examined.
Materials & Methods
- Microplate 384 well, small volume, black (Greiner, #784900)
- ThinSeal™ (ExcelScientific Inc., #100-THIN-PLT)
- CLARIOstar® plate reader, BMG LABTECH
- Acylated and biotinylated histone 3 derived peptides (LifeTein)
- Amyloid fibrils2: RCM α-LA (100µM); in 20 mM sodium phosphate pH 7.4, 100 mM KCl, 10 mM MgCl2, 37°C, slow shaking. RCM ĸ-casein (50 μM); 20 mM sodium phosphate pH 7.4, 37 °C, 300 rpm. Insulin (200 μM); 50 mM glycine-HCl pH 2.0, 60 °C
- Dye concentration: 20µM; total volume: 20µL.
Experimental procedure
For FP experiments, TPE-TPP was pre-mixed with the proteins in solution prior to incubation at higher temperature. FP was recorded in situ with a CLARIOstar microplate reader and the settings indicated below. The background fluorescence of TPE-TPP in the corresponding buffer solution under the respective protein fibrillation conditions was subtracted, and the change in FP values was expressed in milli-polarization units (∆mP).
Instrument Settings
| Optic settings |
Fluorescence Polarisation, plate mode kinetic |
|
| Filters | Ex: 360-20 Dichroic: LP426 Em: 482-16 |
|
| Gain | Channel A: 676 Channel B: 675 |
|
| General settings | Flashes | 50 |
| Settling time | 0.1 s |
|
| Kinetic settings | Cycles | 770 |
| Cycle time | 120s |
|
| Incubation | α-LA, k-casein 37 °C; insulin 60 °C |
|
| Shaking |
α-LA: 100 rpm idle movement k-casein: 300 rpm idle movement |
|
Results & Discussion
TPE-TPP exhibits emission characteristics that are aggregation-specific: it does not give a signal with stable molten globule states or amorphously aggregating species1. The compound can be used to monitor fibril aggregation for various aggregating proteins under various conditions such as; acidic pH, elevated temperature and in the presence of amyloid inhibitors.1 Importantly, TPE-TPP can be used to monitor prefibrillar species via standard fluorescence (data not shown) and FP.
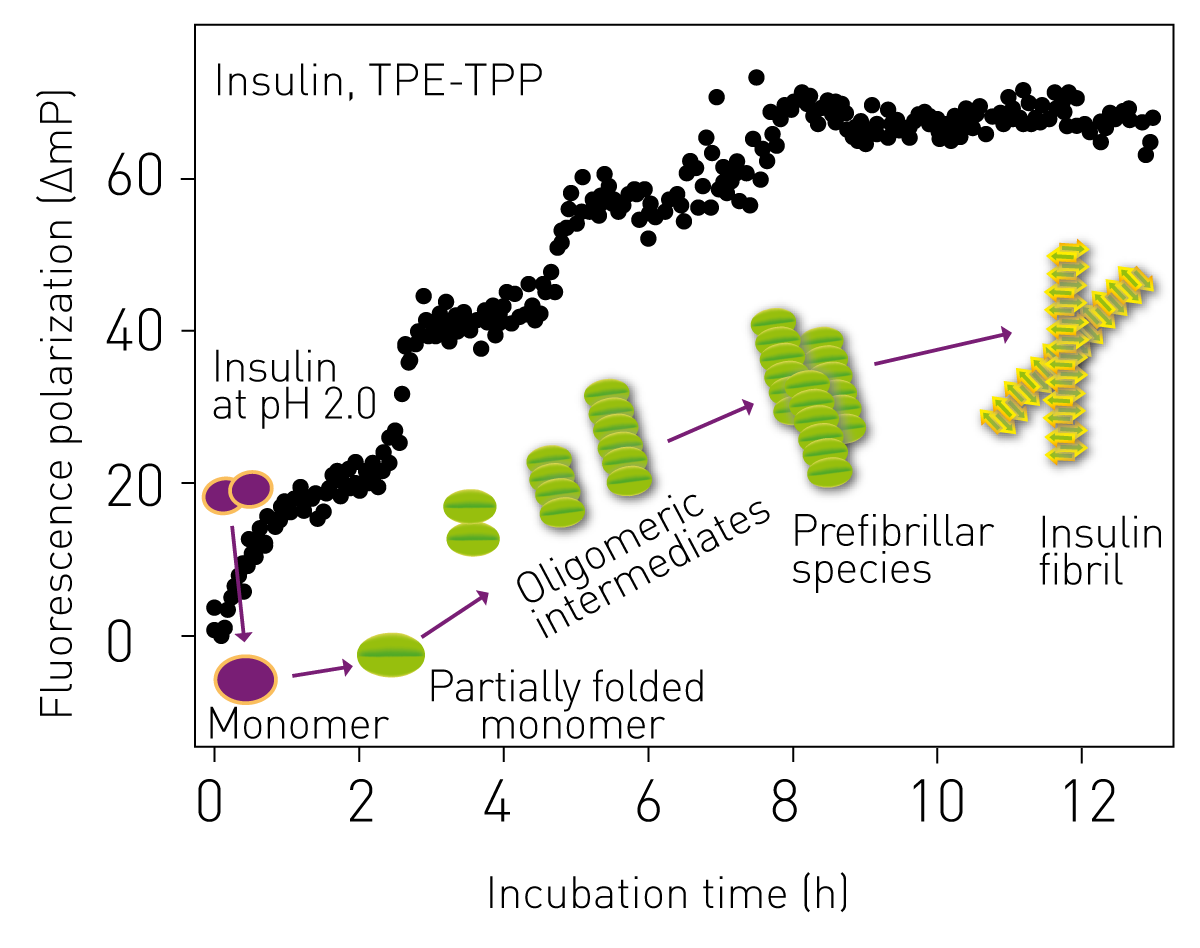
Bovine insulin (monomeric mass 5.8 kDa) forms amyloid fibrils at acidic pH (pH 2.0) and elevated temperature (60°C). Insulin exists as a dimer at acidic pH, which rapidly dissociates to monomer at higher temperature followed by a conformational change. The partially unfolded monomers associate with each other to form oligomeric intermediates including dimers, tetramers and hexamers.3 The FP study of the oligomerisation of monomers of bovine insulin, using TPE-TPP, clearly shows the progression from monomer to oligomers and then to insulin amyloid fibril by the stepwise increase, each of around similar magnitude, in FP value (Fig 2).
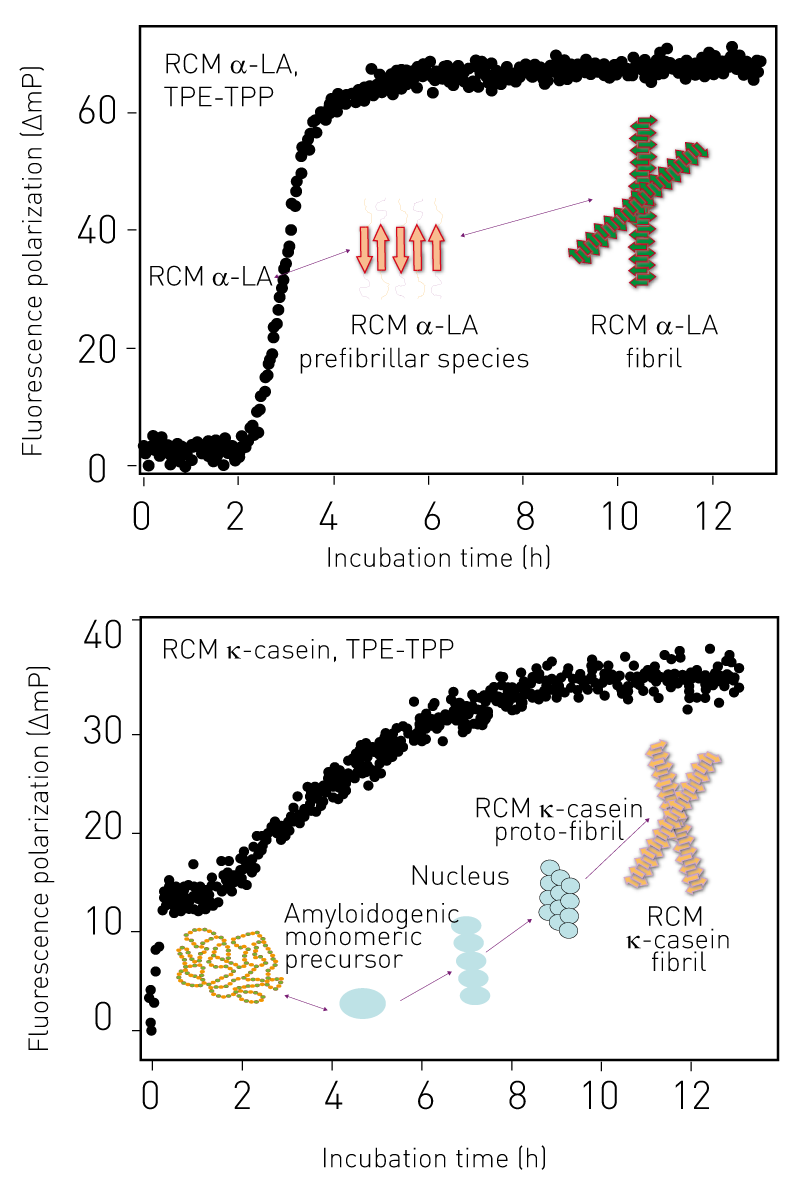
RCM α-LA (monomeric mass 14.7 kDa) follows classical nucleation-dependent polymerization as apparent from its sigmoidal kinetic fluorescence curve.4 Starting from monomers, soluble oligomers with increasing mass were formed after 2 h of incubation, as reflected by the steep increase of FP values. Rigid insoluble fibrils were subsequently formed with FP reaching a maximum value and staying constant over the next 8 h (Fig 3a).
In contrast, RCM ĸ-casein (monomeric mass ~19 kDa) forms amyloid fibrils via a different mechanism with the absence of a significant lag phase. In solution at 25°C, RCM ĸ-casein occurs as a spherical particle with a weight average molecular mass of ~1180 kDa. Consistent with the difference in molecular mass, the FP of RCM ĸ-casein prior to incubation was higher than that of RCM α-LA (Fig 3b). At 37°C and neutral pH, RCM ĸ-casein assembles into fibrillar structures.5 Amyloid fibril formation by RCM ĸ-casein involves the dissociation of its spherical, oligomeric form into a monomeric amyloidogenic precursor, which then undergoes rapid aggregation to form a nucleus from which rigid, rod-like amyloid fibrils develop.6
Conclusion
The results above clearly show that TPE-TPP-based FP measurements can monitor the fibrillar assembly of a variety of amyloid fibril-forming proteins in situ. In addition, the CLARIOstar multi-mode reader (BMG Labtech) is a versatile tool for these studies, offering temperature control to 65°C, heavy duty shaking, and high FP sensitivity.
References
- Kumar, M. Hong, Y. et. al. Anal. Chem. 2017, 89, 9322–9329.
- Kumar,M. Hong,Y.et al.1
- Bekard, I. B.; Dunstan, D. E. Biophys. J. 2009, 97, 2521−2531.
- Kulig, M.; Ecroyd, H. Biochem. J. 2012, 448, 343−352.
- Thorn, D. C. et al. Biochemistry 2005, 44, 17027-17036.
- Ecroyd, H. et al. J. Biol. Chem. 2008, 283, 9012-9022.