Introduction
Measurement of the mitochondrial membrane potential is useful in a wide variety of research areas and mitochondrial dysfunction is implicated in diseases such as cancer, diabetes, Parkinson’s disease, and stroke.
In most eukaryotic cells the majority of ATP production is via oxidative phosphorylation by the respiratory chain. In this way, sugars, such as glucose, and free fatty acids are oxidised, resulting in the pumping of protons across the inner mitochondrial membrane, creating an electro-chemical gradient (the mitochondrial membrane potential). This is in turn used by complex V of the respiratory chain to generate ATP. Therefore the mitochondrial membrane potential makes up a large part of the bioenergetic state of the cell and it is changed directly depending on the cells energy needs. For example fast growing tumour cells have a much higher mitochondrial membrane potential than WT cells, and in turn quiescent or differentiated cells have a still lower membrane potential.
The ability to accurately measure mitochondrial membrane potential can give invaluable information about the general health and function of the mitochondria, in particular of the overall function of the respiratory chain and the potential of the mitochondria to generate ATP and provide energy for other cellular components.
The number of factors which can influence this potential are vast and include many cellular components outside the mitochondria, for example interactions with the activity of the proteasome. In addition, the ability of a cell to maintain its mitochondrial membrane potential can mean the difference between survival of the cell, entry into an apoptotic cell death or a necrotic cell death process. Intact mitochondrial membrane potential is also required for the cell to enter apoptosis. Mitochondrial membrane potential is usually measured using non-invasive cationic dyes. These dyes are sequestered into the mitochondrial matrix in amounts directly proportional to the membrane potential and can then be measured using standard fluorescent techniques, including fluorescence microscopy, flow cytometry or using a microplate reader for high throughput assays.
BMG LABTECH’s FLUOstar Omega microplate reader was used in a high throughput screening assay measuring mitochondrial membrane potential in human cells. The results presented here were gained using primary human fibroblast cells from controls.
Materials & Methods
- Human fibroblast cells obtained from Coriell Cell Repositories
- Black 96 or 384 well plates with transparent bottom from Greiner
- Tetramethylrhodamine methyl ester (TMRM), Sigma
- Carbonyl cyanide 3-chlorophenylhydrazone (CCCP), Sigma
- Ethidium homodimer fluorescent dye, Invitrogen
- FLUOstar Omega microplate reader, BMG LABTECH
Fibroblasts were plated at 40 % confluency in 96 or 384 well plates in minimal growth medium MEM with 10 % FBS, 100 IU/mL penicillin, 100 μg/mL streptomycin, 1 mM sodium pyruvate, 2 mM L-glutamine, 0.1 mM amino acids, 50 μg/mL uridine and 1 X MEM vitamins.
24 hours later cells were changed into galactose culture medium MEM (without glucose), supplemented with 10 % FBS, 100 UI/mL penicillin, 100 μg/mL streptomycin and 0.9 mg/mL galactose, as described before.
After further 24 hours of growth the mitochondrial membrane potential was measured using the fluorescent dye Tetramethylrhodamine methyl ester (TMRM) as described before. Briefly the dye was loaded onto cells at 150 nM in assay buffer (80 mM NaCl, 75 mM KCl, 25 mM D-glucose, 25 mM HEPES, pH 7.4), placed at 37°C for 5 minutes, washed 4 times in PBS and measured on a FLUOstar Omega microplate reader (Excitation: 544 nm and Emission: 590 nm, bottom reading with 50 flashes per well).
Initially TMRM is loaded into the cell via the plasma membrane and subsequently the TMRM dye is sequestered into the mitochondria as the mitochondrial matrix is the most negatively charged part of the cell. Therefore changes in plasma membrane potential could influence the results obtained using TMRM. In order to control for this, each assay is performed in parallel as above plus 10 μM carbonyl cyanide 3-chlorophenyl-hydrazone (CCCP), which collapses the mitochondrial membrane potential. All data is expressed as the total TMRM fluorescence minus the CCCP treated TMRM fluorescence.
It is important to quantify and control for cell number, especially when comparing results between cell lines, therefore cell number is measured using 1 μM ethidium homodimer fluorescent dye in a parallel plate after freeze thawing (Excitation: 544 nm and Emission: 645 nm, bottom reading with 50 flashes per well).
Results & Discussion
Figure 1 shows the mitochondrial membrane potential in 6 control fibroblast cell lines, each measured in triplicate on separate occasions and showing the lack of variation between control cell lines.
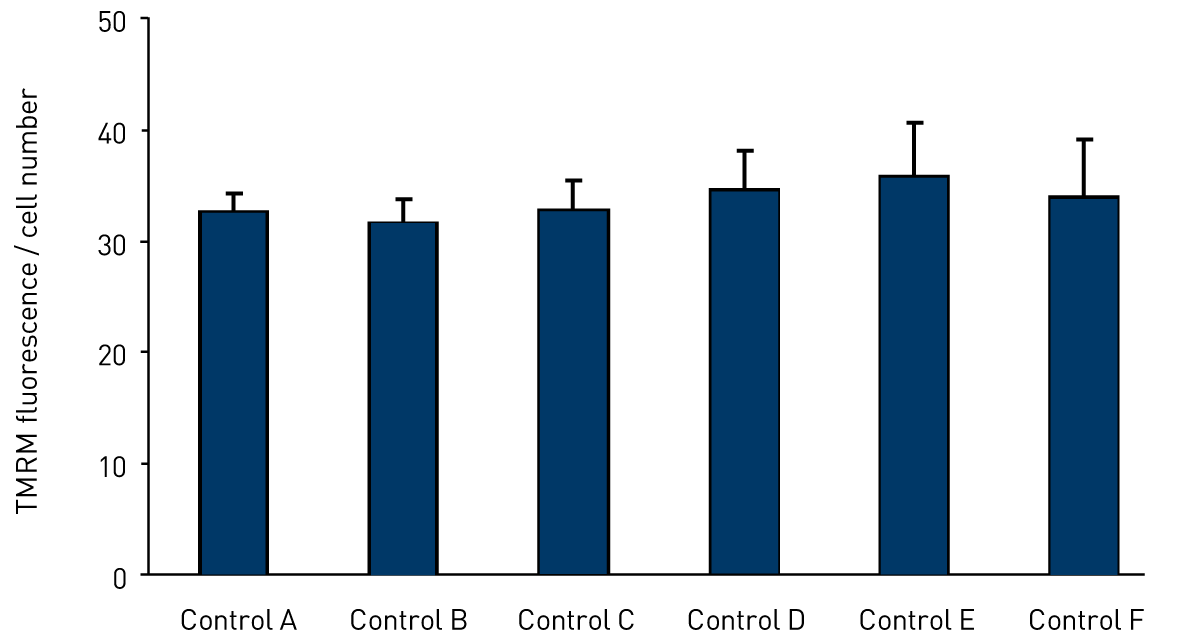 Figure 2 shows the reduction in membrane potential of the same control fibroblasts after treatment with mitochondrial toxin ‘X’ for 72 hours. This demonstrates the range at which the assay is sensitive, creating a large ‘therapeutic window’. Also in this figure the control cells have been treated with an experimental compound ‘Y’ which rescues the reduction in mitochondrial membrane potential caused by treatment with the mitochondrial toxin ‘X’.
Figure 2 shows the reduction in membrane potential of the same control fibroblasts after treatment with mitochondrial toxin ‘X’ for 72 hours. This demonstrates the range at which the assay is sensitive, creating a large ‘therapeutic window’. Also in this figure the control cells have been treated with an experimental compound ‘Y’ which rescues the reduction in mitochondrial membrane potential caused by treatment with the mitochondrial toxin ‘X’.
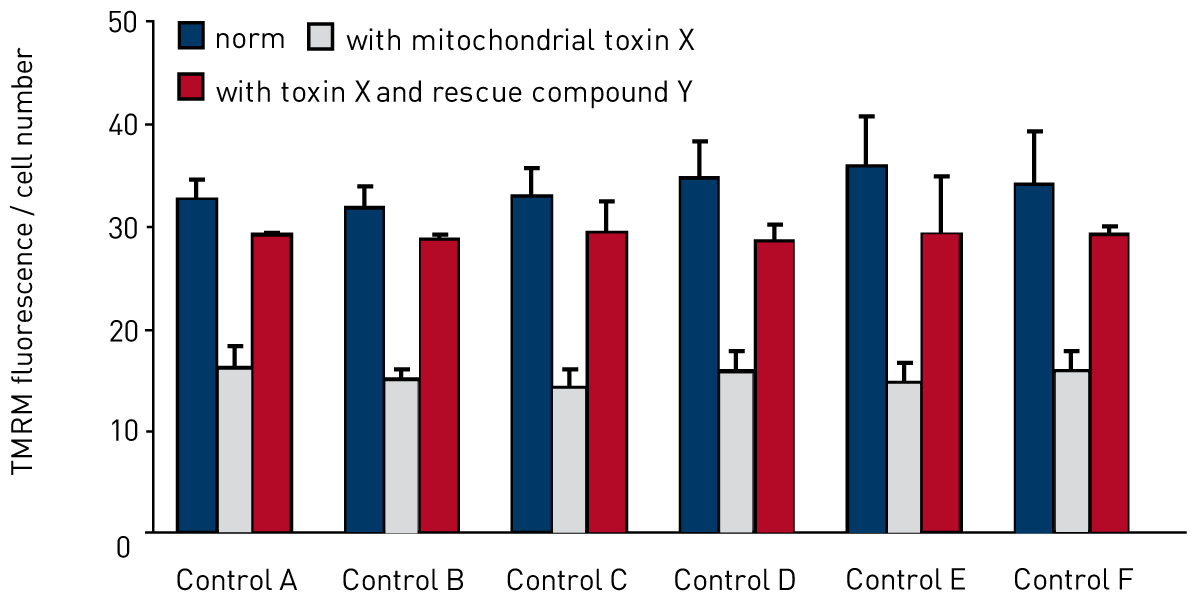
Figure 3 shows that the assay can reliably be scaled up to a 384 well plate format and gain robust, re-producible results.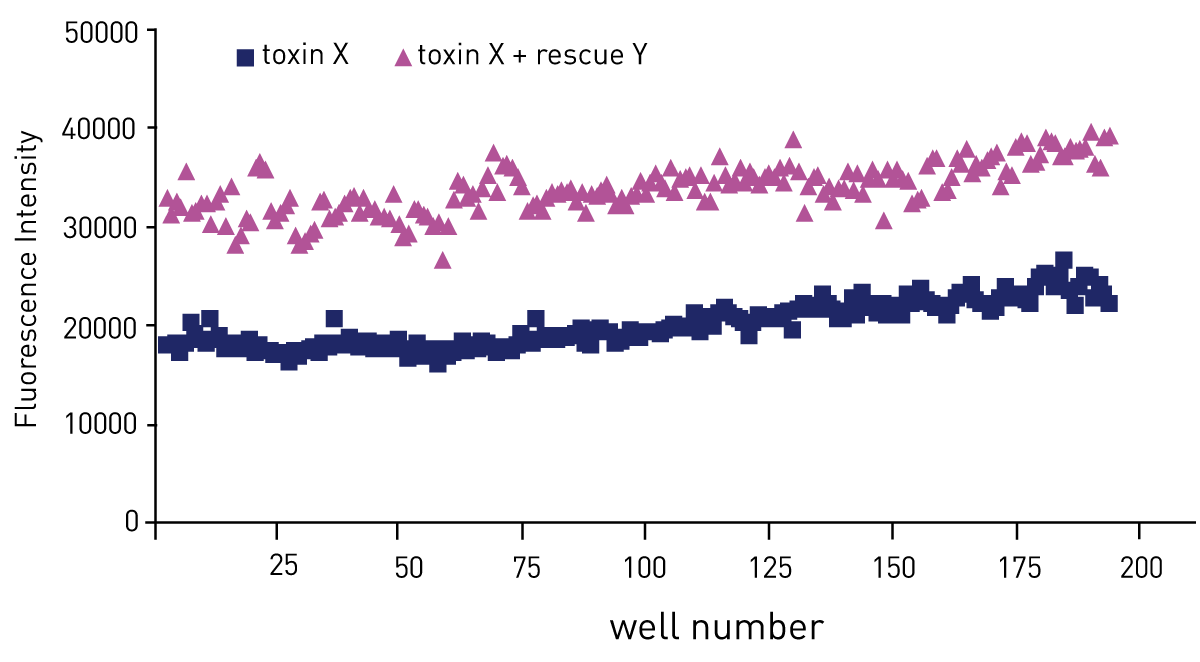
Conclusion
These results show that the FLUOstar Omega is suitable for measuring mitochondrial membrane potential in both 96 and 384 well plate formats. In addition, prolonged low dose treatment of healthy control fibroblast with a mitochondrial toxin causes a significant reduction in mitochondrial membrane potential.
The results are reproducible across a number of different control fibroblast lines and the results of the rescue of this defect with the experimental compound provide further evidence that this assay can be used for high throughput screening using BMG LABTECH’s FLUOstar Omega.