Introduction
Almost all lifeforms exhibit circadian rhythms that repeat approximately every 24 hours and which play an important role in the regulation of biological processes. Circadian rhythms contribute to sleep and feeding patterns in animals and significantly impact health. They also shape the physiological reactions of microorganisms, plants, and animals to the length of day and night. At the heart of these daily rhythms are intricate molecular clocks that convey a sense of time to the function of cells, tissues, and organs. The precise mechanisms by which circadian clocks operate has proven challenging to determine in the complex environment of the cell. Investigations using SDS-polyacrylamide gel electrophoresis reveal the phosphorylation status of protein components of circadian clocks but lack overall sensitivity and do not provide sufficient mechanistic detail for studies of these intricate molecular machines.
In this study, an in vitro clock was reconstituted for Synechococcus elongatus that offers ways to explore how circadian rhythms work in practice and their mechanisms of action.
Assay principle
In vitro fluorescence polarization (FP) can directly monitor circadian rhythms of protein-protein interactions in the cyanobacterial clock in real time when clock proteins are labelled with fluorescent dyes. Free (daytime) and bound (nighttime) clock proteins exhibit significant differences in rotation speeds and the extents of depolarization of the polarized light source. A fluorescence probe assay was developed for each of the individual clock components to permit analysis of the in vitro whole-clock system (Fig. 1).
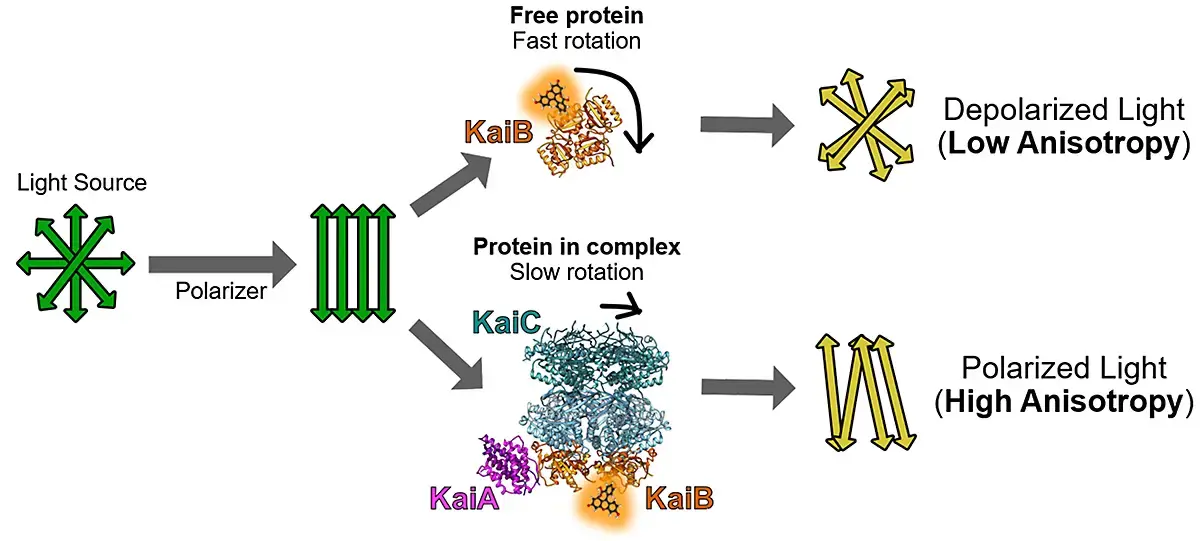
The KaiA, KaiB, SasA, CikA and RpaA protein components were each labeled with the fluorescent probe. The reconstituted system comprised the KaiABC oscillator, SasA and/or CikA, RpaA, and a clock-promoter-bearing DNA fragment (Fig. 2).1 This method can be readily adapted to microplates to enable sensitive fluorescence polarization measurements on a microplate reader.
Materials & methods
-
Black non-binding 384-well microplates (#781900, Greiner BioOne)
-
6-Iodoacetamidofluorescein (#11564896, Invitrogen)
-
CLARIOstar® microplate reader (BMG LABTECH)
Experimental Procedure
Fluorescent probes were attached to clock protein components as described previously using sortase A-mediated ligation.2 Association into larger complexes was monitored using fluorescence polarization. Fluorescently labeled synthetic double-stranded DNA (promoter sequence of kaiBC operon PkaiBC) was used as a representative RpaA target. Each labeled species was monitored in separate parallel reactions. Individual reactions contained 50 to 100 nM of a single fluorescently labeled species for each protein or DNA monitored together with physiological concentrations of each unlabeled clock protein. Plates were read using a BMG LABTECH CLARIOstar® microplate reader to monitor in vitro oscillations in parallel with the following settings:
Instrument Settings
|
Fluorescence polarization, plate mode kinetic
|
||
|
Optic settings |
Filters |
Ex: 482-8 |
|
Gain |
Channel A: 882 |
|
|
Settling time |
0.0 s |
|
|
Kinetic settings |
Number of cycles |
480 |
|
Cycle time |
15 min |
|
|
mP target value |
Set to 10% of maximum theoretical polarization of fluorescein (35 mP) = 35 mP |
|
Fluorescence polarization was converted to fluorescence anisotropy in BMG LABTECH’s MARS software. Fluorescence polarization and anisotropy are mathematically related and used interchangeably.3 All anisotropy values are in mP units.
Results & Discussion
The in vitro clock revealed a set of minimal components to execute clock-controlled gene expression (Fig. 2). This system helped to dissect the mechanistic origins of a clock-disrupting mutant reported in the literature4 and serves as a platform to enable synthetic biology applications. Figure 2 shows fluorescence anisotropy measurements for each clock component in a single microplate and in real time over a duration of several days. The data were fitted to a damped cosine function to extract the period and phase relative to maximal KaiC phosphorylation by reference to its maximum value at circadian time 14 in vivo.4 Circadian times 0 and 12 represent subjective dawn and dusk, respectively, under constant laboratory conditions. The figure shows a comprehensive snapshot of a fully reconstituted in vitro clock complemented by traditional time courses of RpaA and KaiC rhythms of phosphorylation measured in parallel by gel electrophoresis.
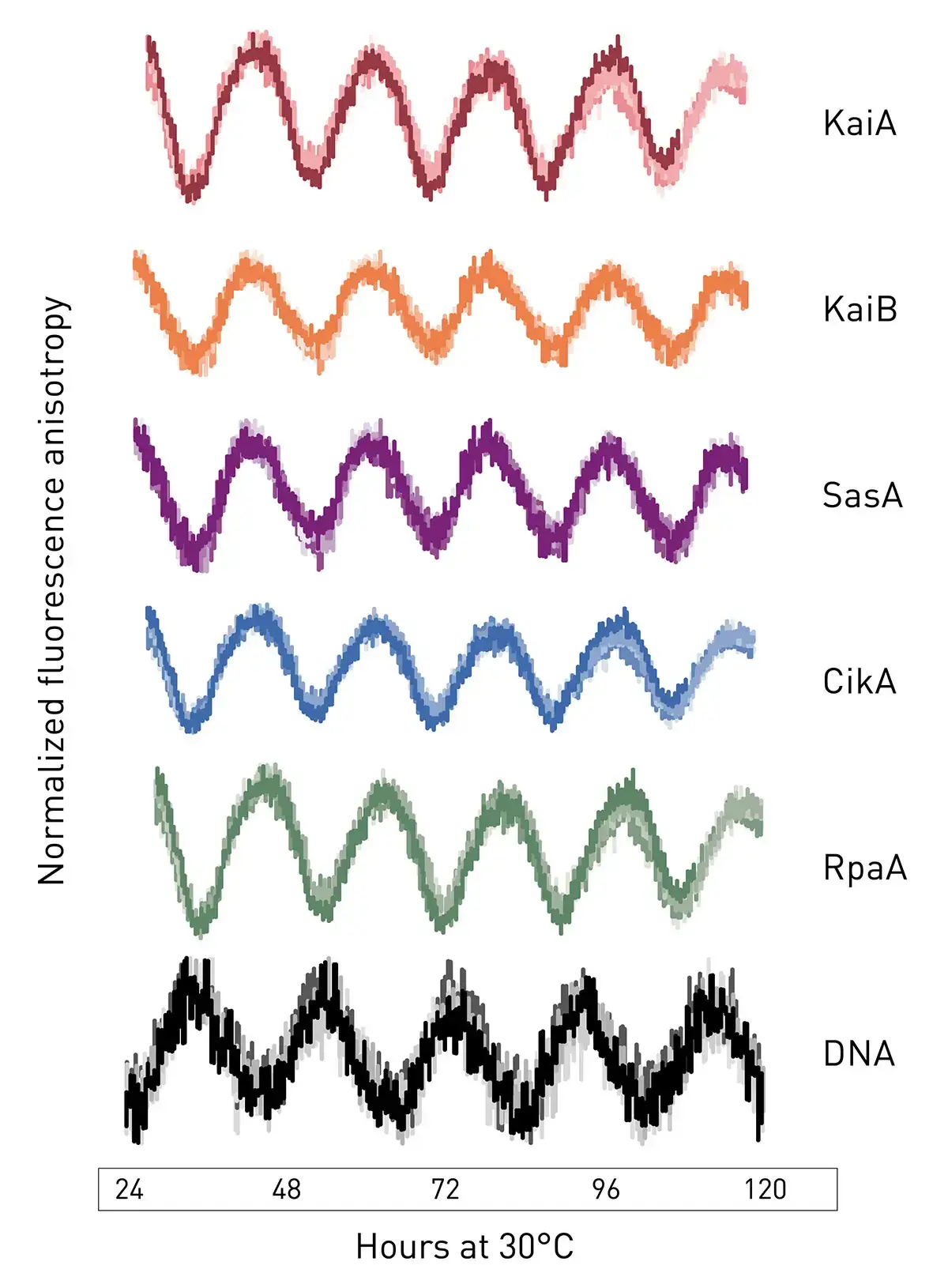
The extended in vitro clock is a free-standing oscillating system whose phases remain coherent for several days without user intervention. The system allowed for observations in real time of the rhythmic behaviors of several clock components in parallel and rhythmically activate the interaction of a transcription factor with a promoter element on DNA. The utility of the system to answer questions about how downstream signal transduction components are coupled to the KaiABC oscillator was investigated. As an example, experiments were performed to determine the mechanistic origin of clock disruption in arrhythmic S. elongatus mutant RpaA-R121Q. This mutant was originally thought to be a crm1 transposon-insertion mutant.5 Further analysis revealed the presence of a single nucleotide polymorphism in the rpaA gene that resulted in an arginine to glutamine substitution at position 121 in RpaA.
Reconstruction of the rpaA-R121Q allele in a wild-type reporter strain displayed the same arrhythmic phenotype which confirmed that mutation of rpaA and not crm1 was responsible for disruption of the clock function (Fig. 3)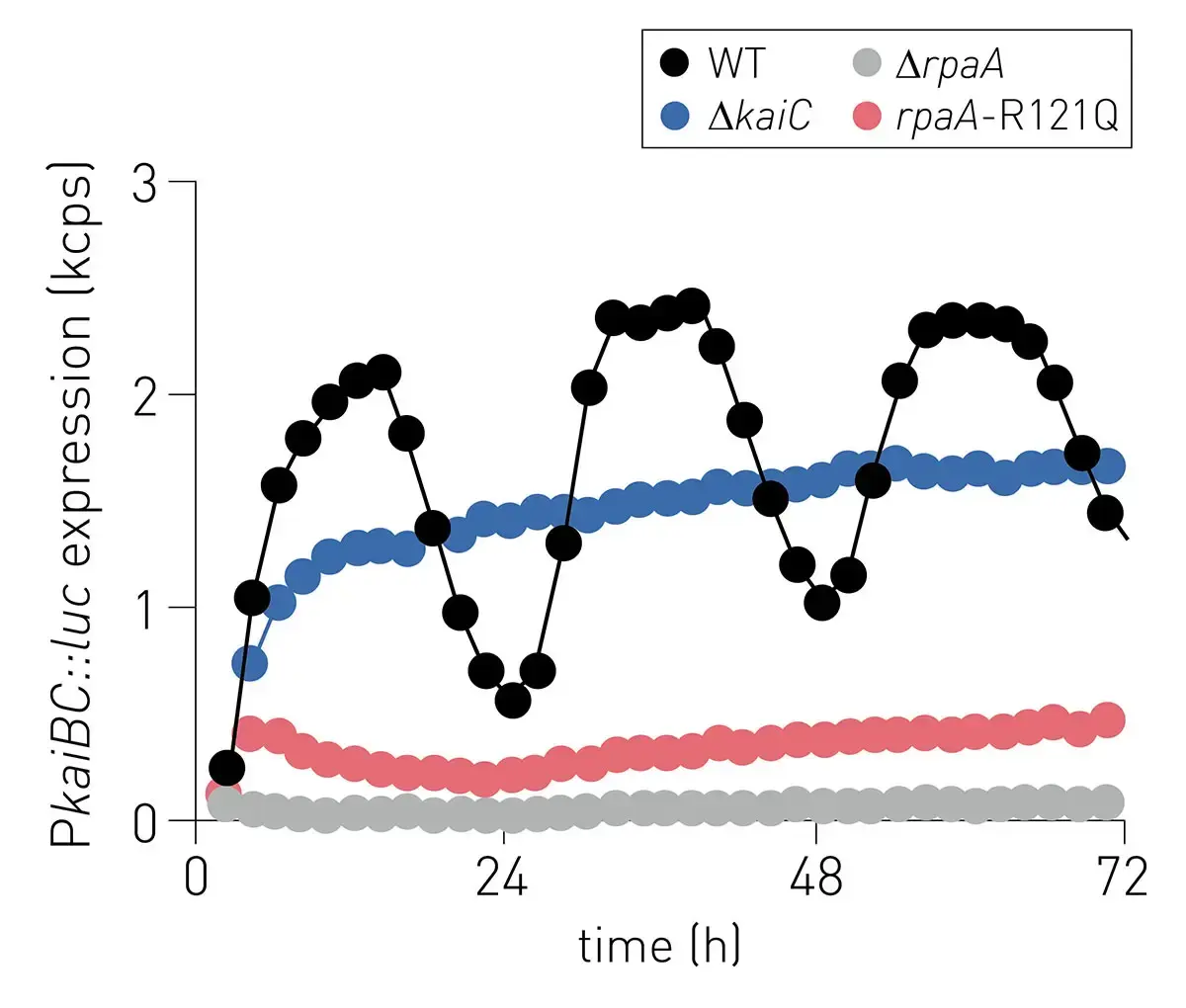
Conclusion
The fluorescence polarization measurements of the in vitro circadian clock in this study allowed hundreds of in vitro reactions to be monitored in real time using a BMG CLARIOstar® microplate reader with a time resolution of minutes. The performance of these assays on a microplate reader delivers significant improvements over conventional analysis by denaturing polyacrylamide gel electrophoresis typically performed after completion of incubations with in vitro clock components. The fluorescence polarization method is not only more sensitive but allows for real time monitoring of all in vitro clock components at scale.
The system serves as a platform for different experimental approaches to study this circadian clock including mathematical modeling related to time control of physiology and metabolism, work in artificial cells, and subsequent engineering of components for synthetic biology applications. It provides an ideal environment to explore the mechanisms involved in the overall operation of this biological clock and its related circadian rhythms.
References
- Chavan, A.G. et al. Science. (2021) 374:(6564): eabd4453.
- Heisler, J. et al. Methods Protoc. (2019) 2(2):42.
- Lakowicz, J.R. Principles of fl uorescence spectroscopy, 3rd edition, Springer: Berlin, Germany, 2006.
- Iwasaki, H. et al. Proceedings of the National Academy of Sciences USA (2002) 99: 15788-15793.
- Boyd, J.S. et al. Proceedings of the National Academy of Sciences USA (2013) 110: 13950-13955.