Introduction
Phagocytosis forms a major part of the innate immune system of higher eukaryotes. It involves the ingestion and destruction of potentially pathogenic or inflammatory particles, such as microorganisms, apoptosing cells, and minerals. In humans, phagocytosis is initiated by the recognition of foreign particles, which can occur via one of two mechanisms. The first involves the direct recognition of repeated patterns of molecular structures on the surface of the microorganisms, such as the Gram-negative bacterial coat protein, lipopolysaccharide (LPS), and heat shock protein-60 (HSP-60).
Alternatively, pathogens can be detected indirectly by the recognition of humoral components from serum, that bind to the surface of pathogens, a process known as opsonisation. The opsonising proteins involved include antibodies and complement proteins which are recognised by Fc and C3b or C3bi receptors respectively. Following recognition, the target is internalised into a phagosome within the cell. This then fuses with a lysosome, which then releases proteases that catalyse the breakdown of the particle.
Alveolar macrophage (AM) phagocytosis of particles and microorganisms is an essential lung host-defence mechanism. In chronic lung diseases, such as chronic obstructive pulmonary disease (COPD), all pathophysiological changes are thought to be mediated by the macrophage. However, studying the process of pathogen clearance in the lung is difficult because obtaining viable AM from bronchoalveolar lavage (BAL) is variable. In contrast, blood cells are easily obtained, therefore a blood macrophage model can be used to examine AM functions. Macrophages are derived from monocytes, which can be separated from peripheral blood mono-nuclear cells (PBMCs). The differentiation of a monocyte to a macrophage takes place over 7-21 days.
Measurement of phagocytosis has traditionally involved microscopy, flow cytometric analysis, or radioactive assays; all of which are time consuming, and in the case of the latter, require additional safety procedures. In this study, the use of a BMG LABTECH microplate reader for quantifying the internalisation of fluorescently labelled phagocytosis targets by monocyte derived macrophages (MDM) was investigated.
The BMG LABTECH readers are able to take multiple readings over time at a user defined temperature and can also be equipped with smart dispensing injection.
Materials & Methods
- BMG LABTECH microplate reader
- Black 96-well clear bottomed culture and assay plates (FisherScientific)
- Fluorescently labelled polystyrene beads (2 μm, carboxylate modified yellow-green, Invitrogen)
- Discontinuous Percoll gradients (Amersham Biosciences)
- Monocyte Isolation Kit II (Miltenyi Biotec)
- Granulocyte macrophage colony stimulating factor (GM-CSF, Biosource)
- Trypan blue (Sigma-Aldrich)
Monocytes were isolated from whole blood obtained from nonsmokers, using dextran sedimentation, discontinuous Percoll gradients and Monocyte Isolation Kit II. These were cultured on black 96-well clear bottomed culture plates with 2ng/ml GM-CSF for 12 days. After this time the monocytes had differentiated to a macrophage-like phenotype.
Alveolar macrophages (AM) were isolated from non-smoking patients undergoing investigative bronchoscopy. Lungs were lavaged with warm saline, which was then retrieved using a fibreoptic bronchoscope. Cells were washed and resuspended in media, then cultured on black 96-well clear bottomed plates. Both kinds of cells, MDM and AM are strongly adherent.
Phagocytosis assays were carried out using fluorescently labelled polystyrene beads (2μm, 50x106 beads/ml). Macrophages (MDM or AM) were incubated with beads for 4h then washed to remove unbound particles. Extracellular fluorescence was then quenched using Trypan blue. The fluorescence of internalised beads was measured on the BMG LABTECH microplate reader with excitation at 480 nm and emission at 520 nm. To confirm internalisation of particles, cells were also viewed using a confocal microscope. DAPI was used to stain nuclei and Evans blue for cytoplasm.
Results & Discussion
In order to establish the optimal incubation time for phagocytosis assays, MDM were isolated incubated for 2, 4 and 6h with fluorescent polystyrene beads. Fluorescence of ingested beads was quantified using a BMG LABTECH microplate reader. These experiments demonstrate that MDM can phagocytose fluorescently labelled polystyrene beads in a time dependent manner (Fig. 1).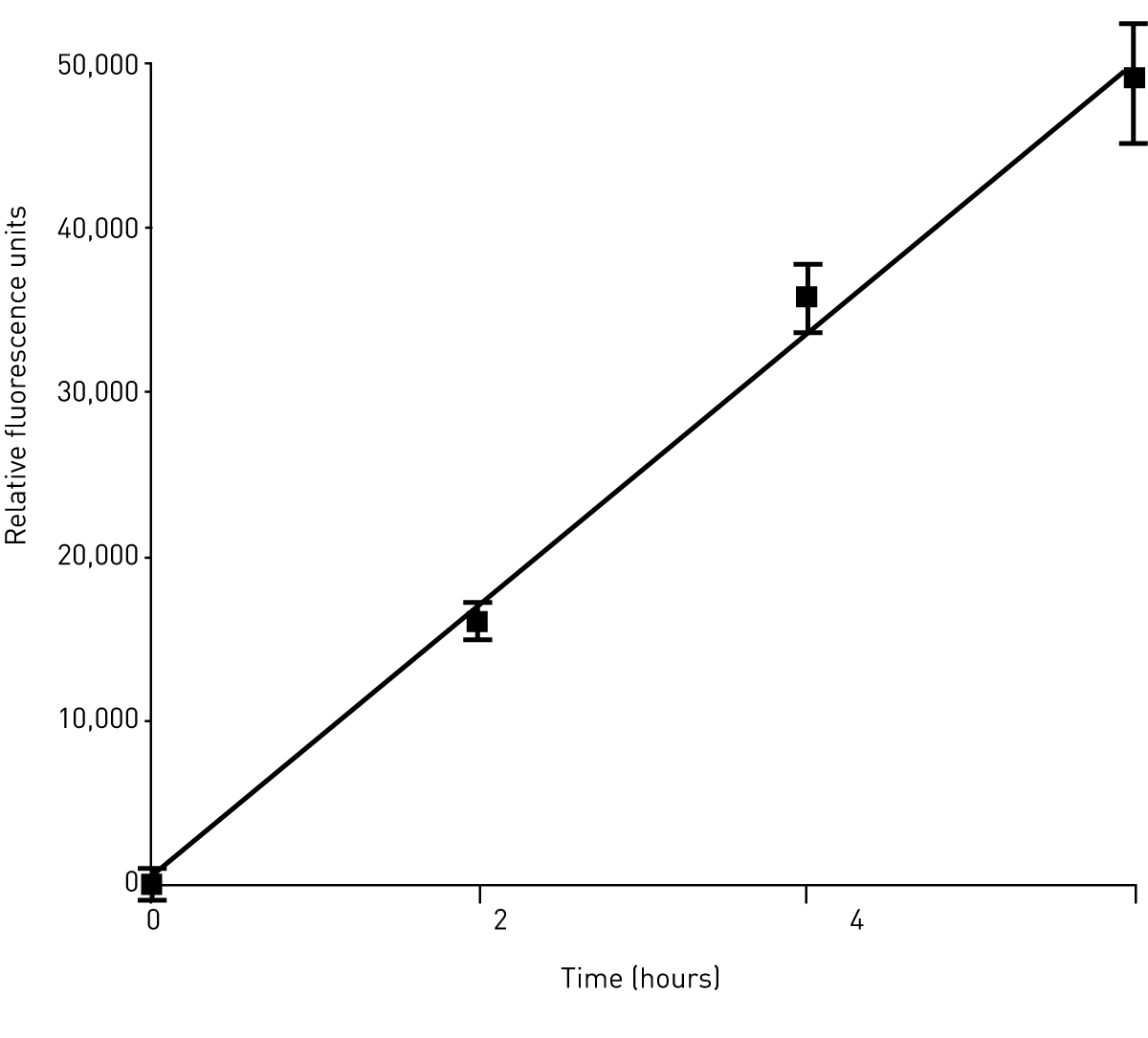
In order to ascertain the optimal concentration of beads to be used in phagocytosis assays, MDM were incubated with solutions of between 0.5-50x106 beads/ml. Phagocytosis of fluorescently labelled polystyrene beads increased in a concentration dependent manner (Fig. 2).
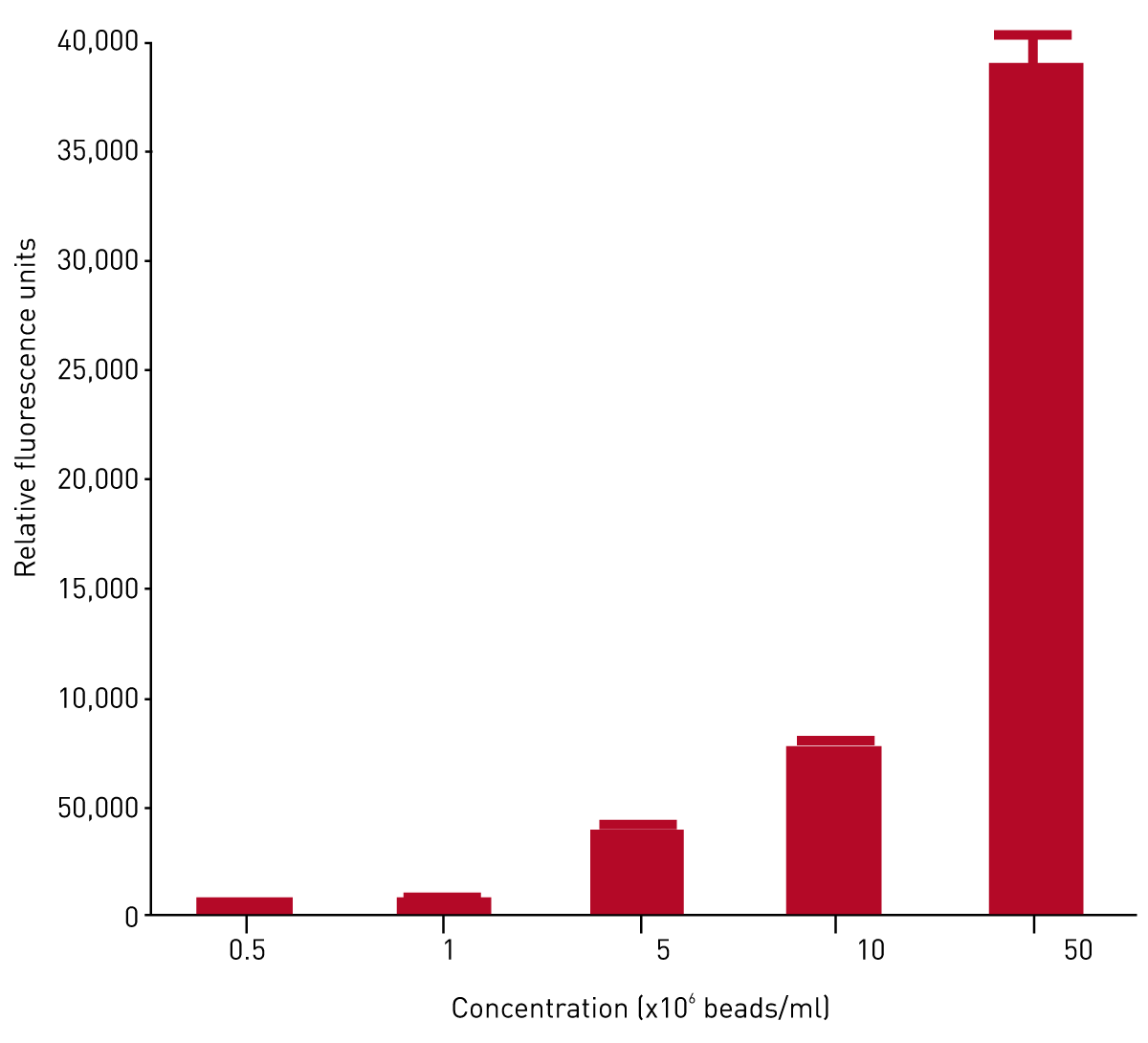
In order to confirm that beads were being internalised and not adhering to cell surfaces, MDM were cultured on chamber slides and incubated with polystyrene beads at increasing concentrations. These slides were then viewed using a confocal microscope. Images taken sequentially through the cells showed the beads to be completely within the cell cytoplasm (images not shown). Additionally, the images verified the dose dependent increase in phagocytosis with concentration of beads. To ensure that MDM phagocytosis reflected that seen in lung derived macrophages, comparative experiments using alveolar macrophages (AM) were performed. MDM and AM were incubated with beads and phagocytosis was measured using a BMG LABTECH microplate reader (figure 3).
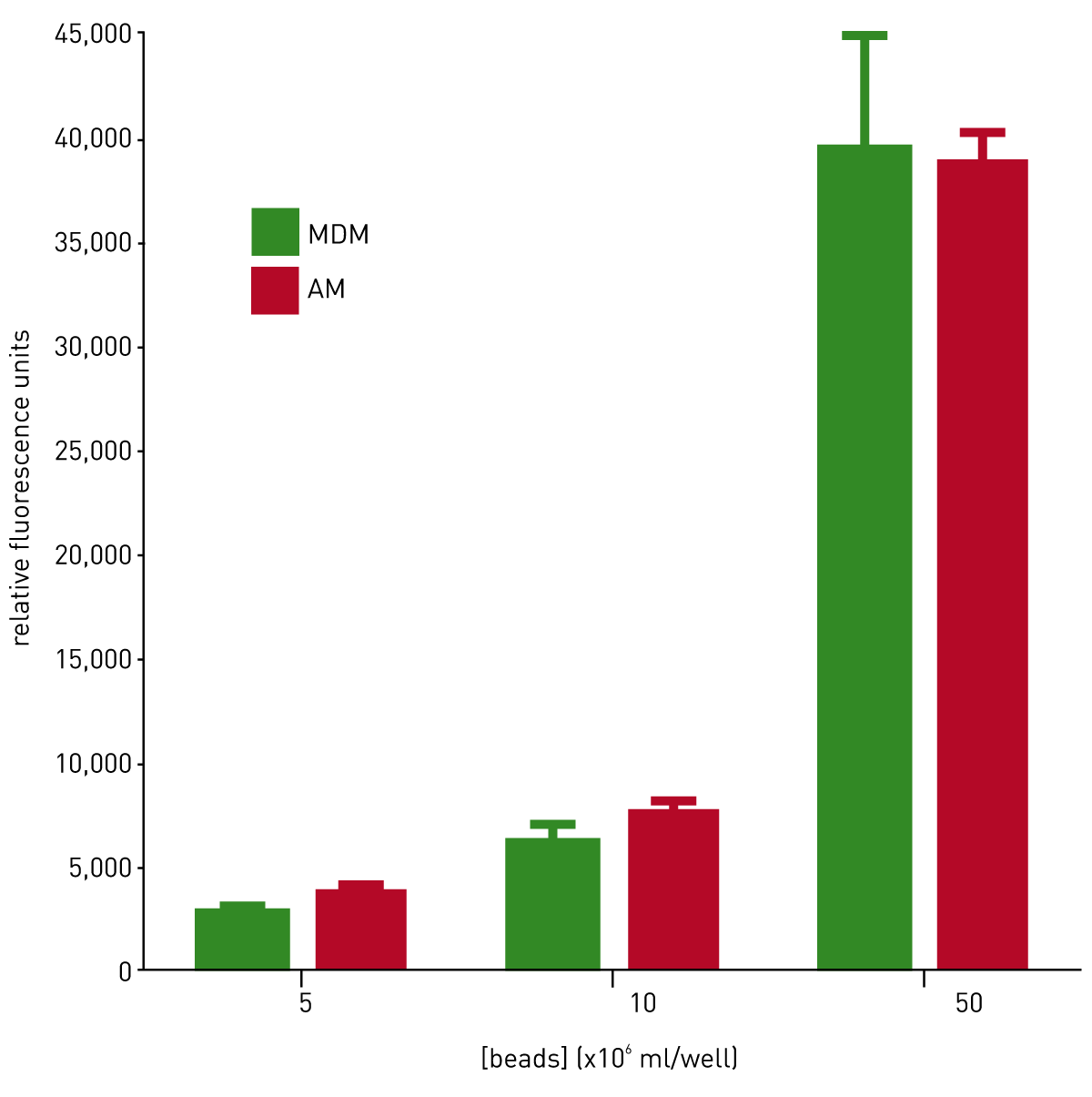
Conclusion
The findings presented here suggest that MDM can phagocytose polystyrene beads in a comparable manner to alveolar macrophages, and therefore are a suitable model to study phagocytosis. The BMG LABTECH microplate reader provides an accurate, consistent method for quantifying macrophage phagocytosis that is significantly more time efficient than previously employed methods.