Introduction
G protein-coupled receptors (GPCRs) are membrane-spanning proteins that transmit extracellular stimuli to the inside of a cell. Activation by stimulating molecules (e.g. neurotransmitter, hormone or chemokine) starts a signaling cascade that results in a cellular response. An example is the chemokine CXCL12 that activates the receptor CXCR4, resulting in G protein activation and recruitment of β-arrestin2. Subsequently, the receptor is internalized for either recycling to the plasma membrane or lysosomal breakdown.
GPCRs are important drug targets requiring receptor-protein interaction and trafficking studies to reveal how they function. Bioluminescence resonance energy transfer (BRET) is a versatile tool to study such interactions and trafficking. However it is limited by the ectopic expression of labelled interaction partners. CRISPR/Cas9 genome editing overcomes the limitation by enabling endogenous expression of luciferase-labelled proteins.
Assay Principle
BRET uses a luciferase that in the presence of a substrate produces blue light. If a suitable fluorophore is in very close proximity and appropriate orientation, less light is emitted and resonance energy is transferred. Thus, interaction of fluorophore- and luciferase-labelled proteins can be monitored by relating fluorophore emission to luciferase signal.
The CRISPR/Cas9 genome editing method incises dsDNA at a precise target site. In the presence of a donor DNA template, homology-directed repair results in the insertion of the donor sequence.
CRISPR/Cas9 enabled insertion of the DNA coding for nanoluciferase (Nluc) into the endogenous genomic locus of CXCR4 of HEK293FT cells. The resulting CXCR4/Nluc fusion protein acts as a BRET donor and overcomes the need for exogenous donor expression.
Materials & Methods
- White 96-well plate (Greiner)
- CLARIOstar® and LUMIstar® (BMG LABTECH)
- CXCL12 (Preprotech), AMD3100 (Sigma-Aldrich), Isoproterenol (Sigma-Aldrich)
- Furimazine (Promega)
Experimental procedure
For detailed descriptions please refer to White et al. (2017). Briefly, CRISPR/Cas9 genome-edited HEK293FT cells expressing CXCR4/Nluc were transfected with cDNA coding for the protein with acceptor-fluorophore using Fugene®6 (Promega). 48 h after transfection cells were incubated with the luciferase substrate furimazine and filtered light emissions were analyzed on a CLARIOstar or LUMIstar Omega.
| Settings stable through experiments | ||
| Luminescence, top optic, plate mode | ||
| Measurement interval time 1 s | ||
| Incubation at 37 °C | ||
| Optic settings (Filters or monochromator and [gains]) | ||
| Fig. 1 | Fig. 2 | Fig. 3 |
| CLARIOstar Filters Nluc 450-60 (3400) Venus 570-100 (2800) |
CLARIOstar Monochromator Nluc 450-60 (3600) Venus 550-60 (3600) HaloTag 660-100 (3200) |
LUMIstar Filters Nluc 475-30 (3200) Venus 535-30 (3600) |
| Number of cycles / Cycle time |
||
| 40 / 33 s | 30 / 49 s | 60 / 31 s |
Results & Discussion
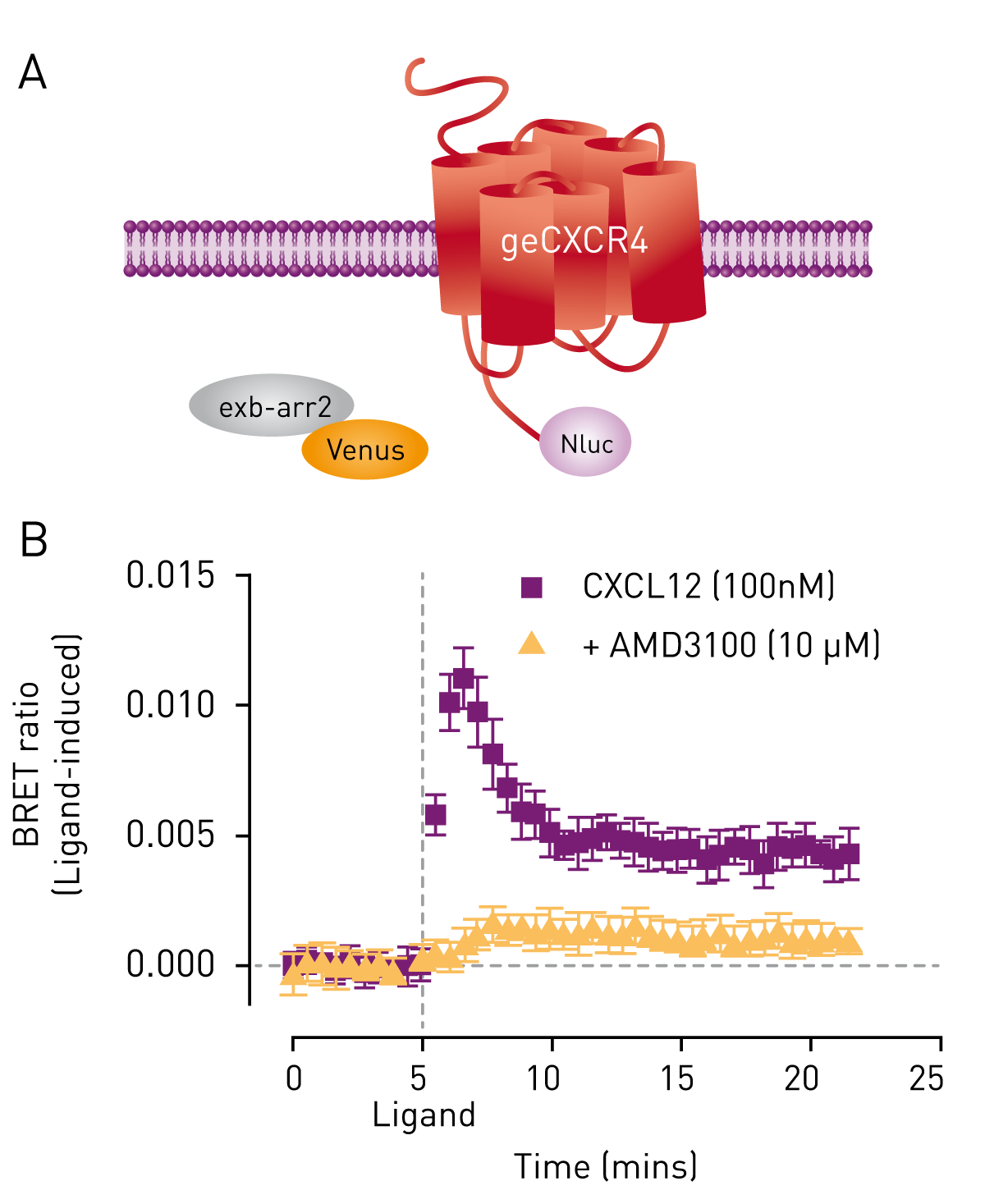
The suitability of genome-edited cells expressing CXCR4/Nluc to report on receptor interaction was studied by combining it with exogenously expressed β-arrestin- 2/Venus (Fig. 1A). Upon activation of CXCR4 with its ligand CXCL12, β-arrestin2 was recruited as reported by an increase in the BRET ratio (Fig. 1B, purple). The CXCR4 antagonist AMD3100 inhibited CXCL12-induced recruitment of β-arrestin2 as seen by the reduction in energy transfer between CXCR4/Nluc and β-arrestin2/Venus (Fig. 1B, orange).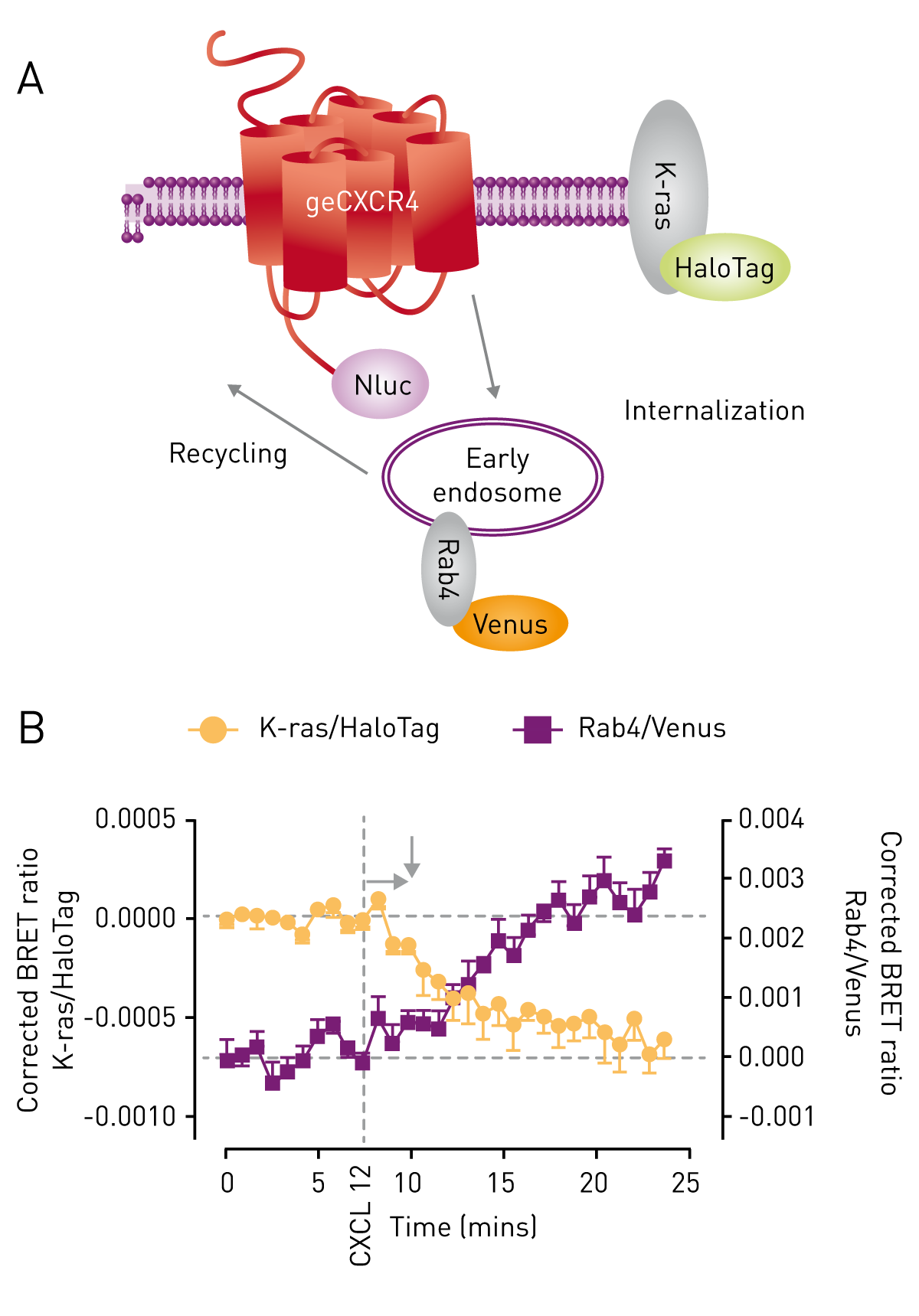 Genome-edited cells endogenously expressing CXCR4/Nluc were further used to study receptor internalization and trafficking. To this end, cells were transiently co-transfected with a K-ras fragment fused to HaloTag and a Rab4-Venus fusion protein serving as plasma membrane and early endosome marker, respectively (Fig. 2A). As signals of Venus and HaloTag can be separated using the CLARIOstar’s monochromator, BRET ratios of the respective proteins were determined in one experiment. Upon addition of the agonist, CXCR4 dissociates from the plasma membrane reported by a decrease in BRET ratio of the K-ras marker (Fig. 2B, orange). At the same time, an increase in the BRET ratio and therefore proximity of the receptor and Rab4 is observed (Fig 2B, purple). The data indicate that the receptor is shuttled from the plasma membrane to the early endosome.
Genome-edited cells endogenously expressing CXCR4/Nluc were further used to study receptor internalization and trafficking. To this end, cells were transiently co-transfected with a K-ras fragment fused to HaloTag and a Rab4-Venus fusion protein serving as plasma membrane and early endosome marker, respectively (Fig. 2A). As signals of Venus and HaloTag can be separated using the CLARIOstar’s monochromator, BRET ratios of the respective proteins were determined in one experiment. Upon addition of the agonist, CXCR4 dissociates from the plasma membrane reported by a decrease in BRET ratio of the K-ras marker (Fig. 2B, orange). At the same time, an increase in the BRET ratio and therefore proximity of the receptor and Rab4 is observed (Fig 2B, purple). The data indicate that the receptor is shuttled from the plasma membrane to the early endosome.
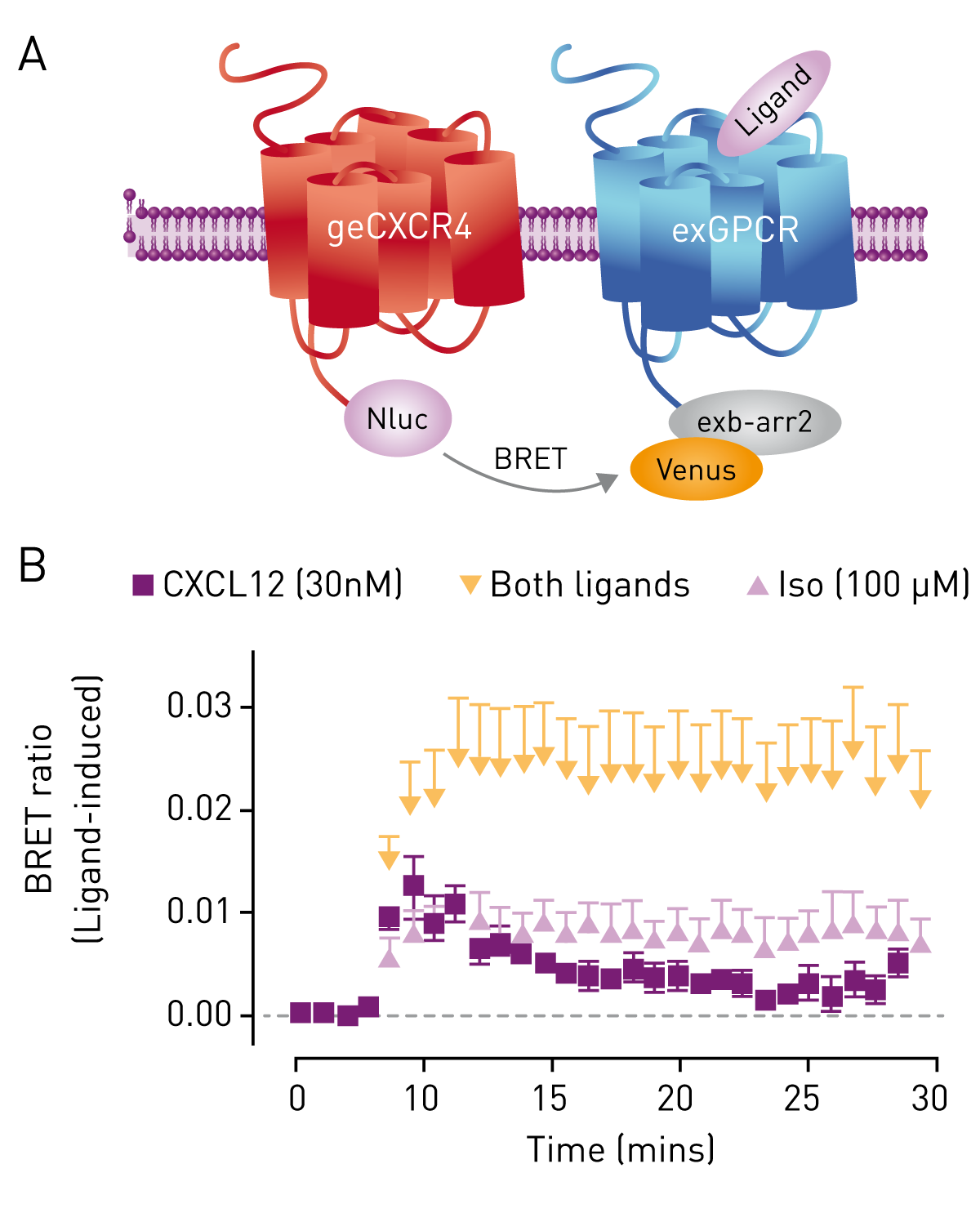
The GPCR Heteromer Investigation Technology (GPCR-HIT, Dimerix) reports on GPCR heteromer formation by the recruitment of an interacting protein specifically to the heteromer complex. Cells expressing CXCR4/Nluc were transiently transfected with cDNA coding for β2-adrenoceptor as well as the interacting protein β-arrestin2/Venus. Treating the cells with CXCL12 resulted in the expected recruitment of β-arrestin2/Venus to genome-edited CXCR4/Nluc, as did treatment with the β2-adrenoceptor agonist isoprenaline, indicating the close proximity of β2-adrenoceptor to CXCR4/Nluc. Applying both agonists resulted in a greater than additive BRET signal, again suggestive of heteromer formation.
Conclusion
The novel CRISPR/Cas9 technique successfully fused the Nluc BRET donor to endogenously-expressed CXCR4. Luminescence generated by the resulting protein-luciferase fusion was sufficient to monitor receptor-protein interactions as well as trafficking. The multiplex internalization assay depends on two acceptor fluorophores whose selective detection was rendered possible by the CLARIOstar’s monochromator.
References
- White CW et al. (2017) Sci Rep. 7:3187. doi: 10.1038/s41598-017-03486-2