Introduction
Protein acetylation is a universal regulatory mechanism of protein activities. Recent developments in epigenetic drug discovery have caused increasing interest in methods to study lysine deacetylation, including in vitro assays of lysine deacetylases.1 Here we optimized a commercial peptidase coupled lysine deacetylase assay to establish the enzymatic properties of Escherichia coli CobB. CobB is a sirtuin-type enzyme that participates in the regulation of bacterial transcription, translation, metabolism, and chemotaxis.2-4.
Assay Principle
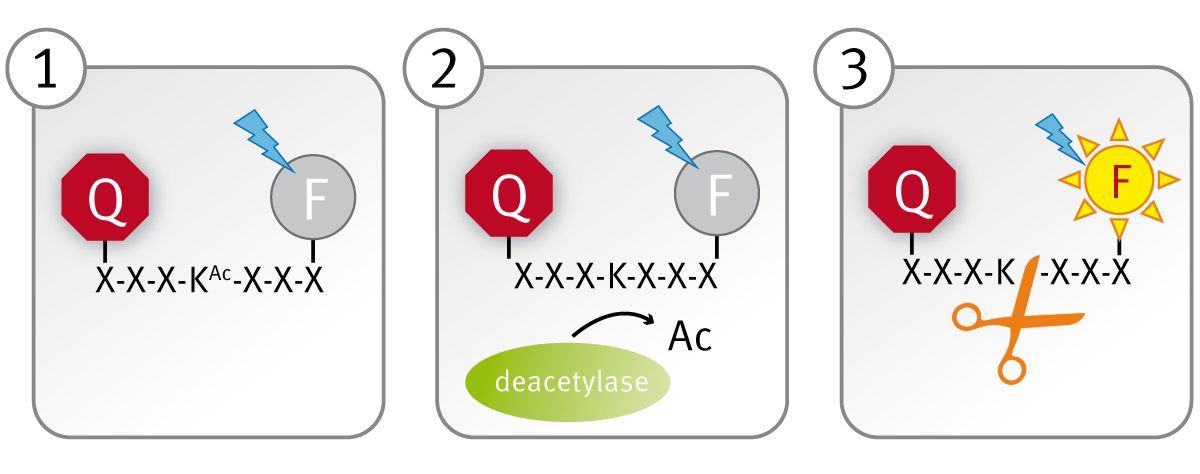 The CycLex SIRT1 assay kit makes use of an acetylated peptide conjugated to both a fluorophore and its quencher (panel 1). Enzymatic deacetylation of a lysine residue within the peptide (panel 2) generates a functional peptidase cleavage site. Fast subsequent cleavage of the deacetylated peptide by the peptidase separates the fluorophore from the quencher and allows fluorescent readout of the reaction (panel 3).
The CycLex SIRT1 assay kit makes use of an acetylated peptide conjugated to both a fluorophore and its quencher (panel 1). Enzymatic deacetylation of a lysine residue within the peptide (panel 2) generates a functional peptidase cleavage site. Fast subsequent cleavage of the deacetylated peptide by the peptidase separates the fluorophore from the quencher and allows fluorescent readout of the reaction (panel 3).
Materials & Methods
- 384-well black flat-bottom microplates (Greiner, #781900)
- SIRT1 fluorimetric assay kit (CycLex, CY-1151V2) Recombinant Escherichia coli CobB
- CLARIOstar microplate reader from BMG LABTECH
All standard chemicals and disposables were obtained through normal distribution channels. His6-tagged E.coli CobB was expressed in E.coli BL21(DE3) cells transfected with a pET28 expression vector, and purified using standard protocols involving immobilized metal ion chromatography (HisTrap, GE Healthcare) and size exclusion chromatography (Superdex-75, GE Healthcare). CobB lysine deacetylase activity was measured at ambient temperature (21°C) under the general conditions described in the manufacturer’s protocol, with modifications as described below. Enzymatic assays were carried out using 50 nM CobB with the addition of NAD+ and inhibitor as indicated.
Instrument settings
| Detection Mode: | Fluorescence Intensity |
| Detection Method | Plate Mode Kinetic |
| No. of cycles: | 120 |
| Cycle time (sec): | 60 |
| No. of flashes: | 5 |
| Scan mode: | orbital averaging |
| Scan diameter (mm): | 1 |
| Excitation: | 355-10 |
| Dichroic: | 400 (autoset) |
| Emission: | 445-10 |
| Shaking: | 8 sec before each cycle |
| Shaking mode: | orbital, 400 rpm |
Results & Discussion
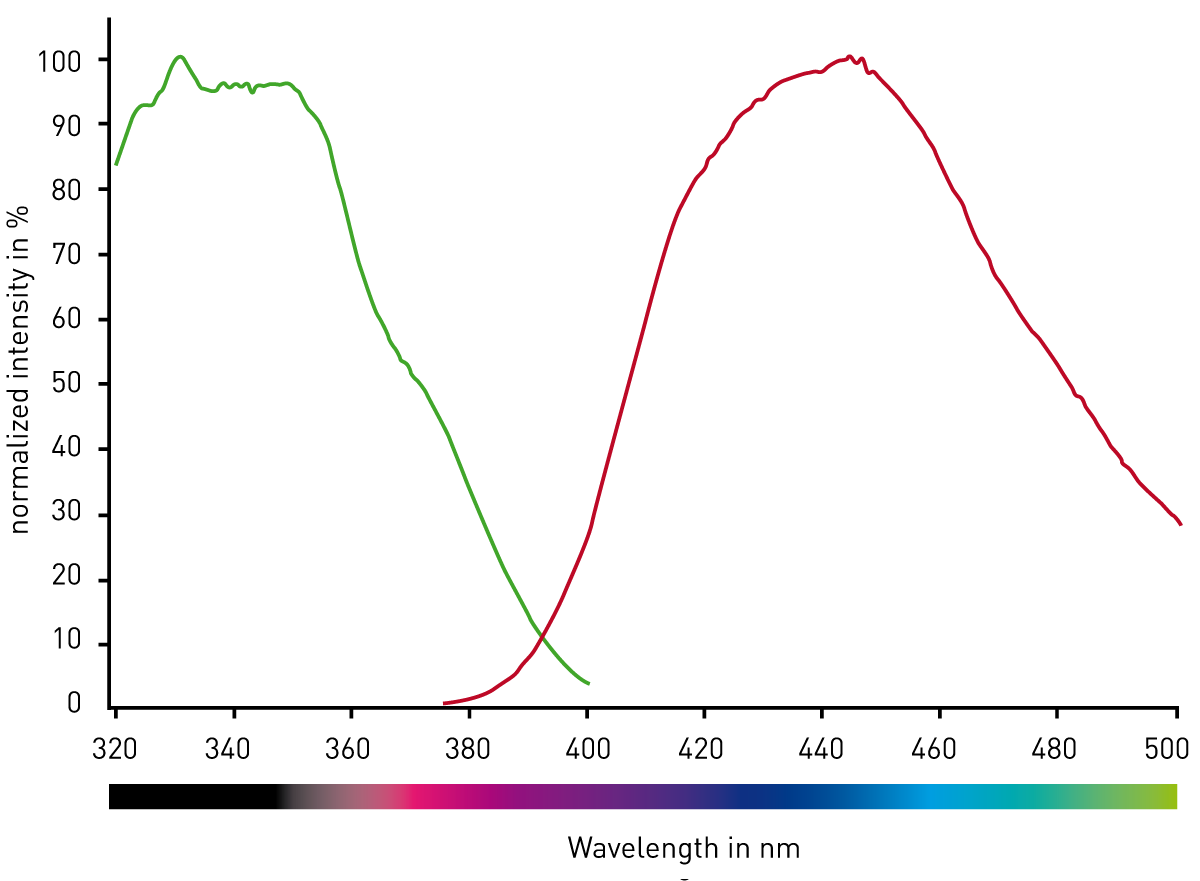
Optimization of excitation and emission parameters Spectral scans (Fig. 2) of the fluorogenic peptide provided with the assay kit indicated that the excitation and emission settings could be improved. We used the CLARIOstar monochromators with excitation at 355/10 nm, a dichroic set at 400 nm, and emission at 445/10 nm. This resulted in the highest signal, although it did not change the Z’ factor5 which was 0.78 using either the above settings or the settings recommended by the provider.
Reference curve for kinetic measurements
A dilution series of the deacetylated reference peptide provided with the assay kit was used to calibrate the measurements (not shown).
Basic kinetic parameters of CobB lysine deacetylase activity
We conducted CobB enzymatic assays in presence of various concentrations of NAD+ and fluorogenic substrate peptide at the concentration recommended in the SIRT1 assay kit manual. Under our experimental conditions, we determined a KMNAD+ of 71 μM (Fig. 3). We are not aware of published kinetic parameters for NAD+ cleavage by CobB.

Inhibition of CobB by a general sirtuin inhibitor
Ex527 is a general inhibitor of sirtuins.6 We determined the potency of Ex527 for the inhibition of CobB activity. Assays were carried out using 70 μM NAD+ as co-substrate and varying concentrations of Ex527 with a final DMSO concentration of 1%. The result of this analysis is shown in Fig. 4.
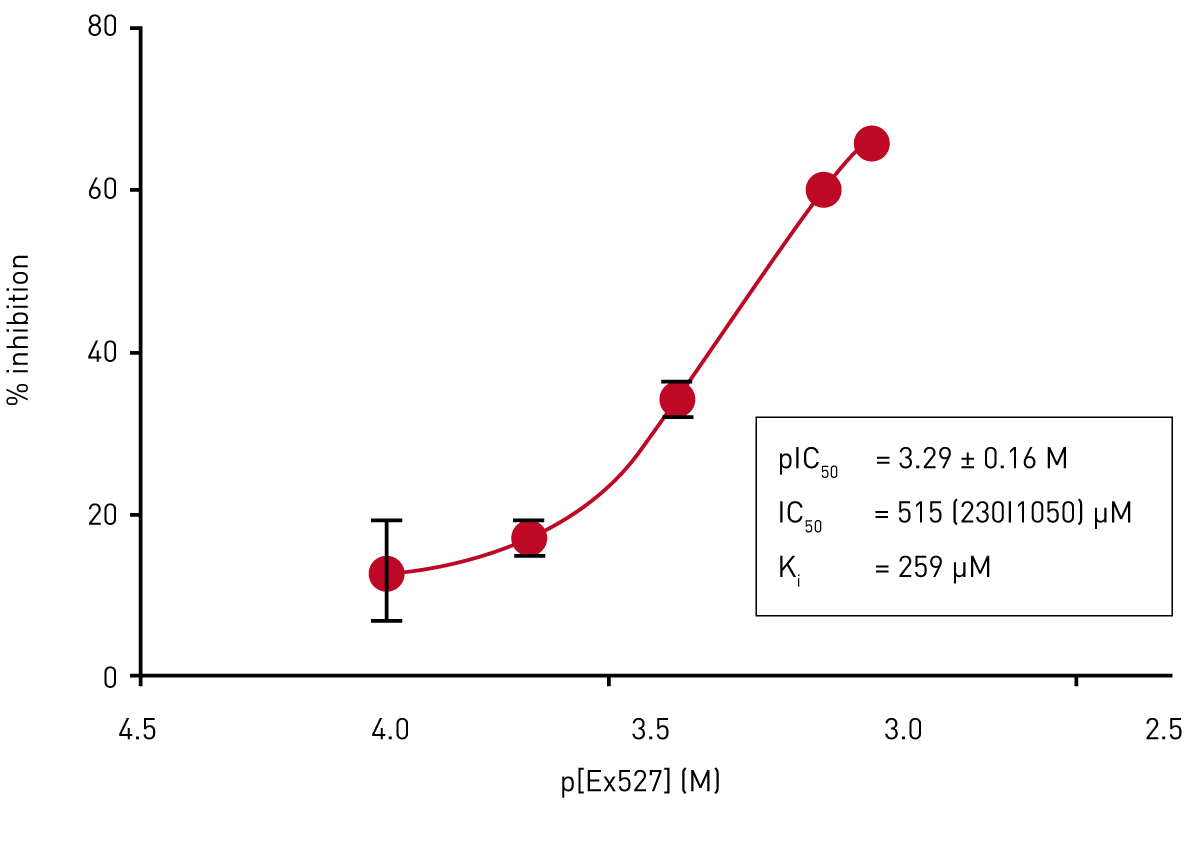 Ex527 is a weak CobB inhibitor (IC50 515 μM; Ki = 259 μM determined using the Cheng-Prusoff equation). The low potency of the inhibitor, in combination with its limited solubility and the DMSO sensitivity of the enzyme, compromised the quality of the determination; the effect of Ex527 concentrations higher than 1 mM could not be measured at 1% DMSO.
Ex527 is a weak CobB inhibitor (IC50 515 μM; Ki = 259 μM determined using the Cheng-Prusoff equation). The low potency of the inhibitor, in combination with its limited solubility and the DMSO sensitivity of the enzyme, compromised the quality of the determination; the effect of Ex527 concentrations higher than 1 mM could not be measured at 1% DMSO.
Conclusion
We have used the CLARIOstar multi-mode microplate reader to determine the kinetic properties of the E.coli lysine deacetylase CobB and its inhibition by a general sirtuin inhibitor, using a commercial assay kit. Our results illustrate the general method, which should be useful for similar investigations into other lysine deacetylases and, in principle, lysine directed acetyltransferases.
Acknowledgments
The CobB expression vector was kindly provided by Dmitry Ivanov (A-Star Institute of Molecular and Cell Biology, Singapore). This work was supported by the Swedish Foundation for Strategic Research, the IngaBritt and Arne Lundbergs Research Foundation, and Karolinska Institutet.
References
- Huston et al. (2015) Nat. Chem. Biol. 11, 542-545.
- Baeza et al. (2014) J. Biol. Chem. 289, 21326-21338.
- Thao et al. (2010) PLoS One.5(12), e15123.
- Li et al. (2010) Mol. Microbiol. (76), 1162-1174.
- Zhang et al. (1999) J. Biomol. Screen. (4), 67-73.
- Ekblad & Schüler (2016) Chem. Biol. Drug Des. 87, 478-482.