Introduction
PARP (Poly(ADP-ribose) polymerase) family enzymes are involved in the regulation of transcription, DNA repair, and chromatin remodeling. These enzymes use nicotinamide adenine dinucleotide (NAD) as a substrate to build poly(ADP-ribose). Due to various links to diseases, PARP enzymes are targets for pharmaceutical drug development.
In this application note, we describe the use of a chemiluminescent assay to determine PARP activity on the CLARIOstar multimode microplate reader. The assay allows kinetic analysis of PARP enzymes and evaluation of inhibitor potency.
Assay Principle
PARP activity is followed in vitro by detecting the incorporation of biotinylated ADP-ribose as a consequence of either enzyme target protein modification or auto-modification. The reaction principle is shown in Fig. 1.
Hexahistidine-tagged PARP enzyme or protein substrate is immobilized on Ni2+-chelating microplates. The reaction is started by adding biotinylated NAD+. The PARP enzyme uses the NAD+ to synthesize bio-tinylated poly(ADP-ribose). This polymer is either added to the PARP enzyme itself or transferred to a protein substrate on the microplate (histone). After a washing step streptavidin-conjugated horseradish peroxidase is added and will bind to the biotinylated poly(ADP-ribose). After adding a substrate to the horseradish peroxidase, chemiluminescence is released and can be measured.
Materials & Methods
- Ni-NTA-coated, opaque, white 96-well microplates (5-PRIME)
- Streptavidin-conjugated horseradish peroxidase (Jackson Immunoresearch)
- Biotinylated NAD+ (Trevigen)
- SuperSignal West Pico chemiluminescent substrate (Thermo Fisher Scientific)
- Microplate reader from BMG LABTECH
All standard chemicals and disposables were obtained through normal distribution channels.
Enzymatic reactions
Hexahistidine-tagged PARP enzyme or protein substrate was immobilized on Ni2+-chelating plates. ADP-ribosylation reactions were started by addition of NAD+ (2 % biotinylated) at 20°C. Reactions were stopped by addition of 7 M guanidine hydrochloride. Plate wells were washed with reaction buffer, incubated for 30 minutes with TRIS-buffered saline containing 0.02 % Tween-20 (TBST) and 1 % (w/v) BSA, and washed with TBST. After incubation with streptavidin-conjugated horseradish peroxidase (0.5 μg/ml) another washing step was done. After adding SuperSignal West (50 + 50 μl, undiluted) as substrate for the peroxidase chemiluminescence was detected in the CLARIOstar microplate reader using the following instrument settings.
Instrument settings
All measurements (linear range check, KM determination, and inhibitor dose-response) were done in endpoint mode.
| Optic: | top optic used |
| Measurement interval time (s): | 1.00 |
| Presetname: | Enliten ATP |
| Emission: | full range (no filter) |
| Gain: | needs to be adjusted prior the measurement |
| Focal height: | needs to be adjusted prior the measurement |
Results & Discussion
For validation of the ADP-ribosyltransferase assay the linear range of signals obtained by a dilution series of biotin-ADP-ribosylated enzyme was determined (Fig. 2). The results show that the signal is linear over a wide range of ADP-ribosyl concentrations.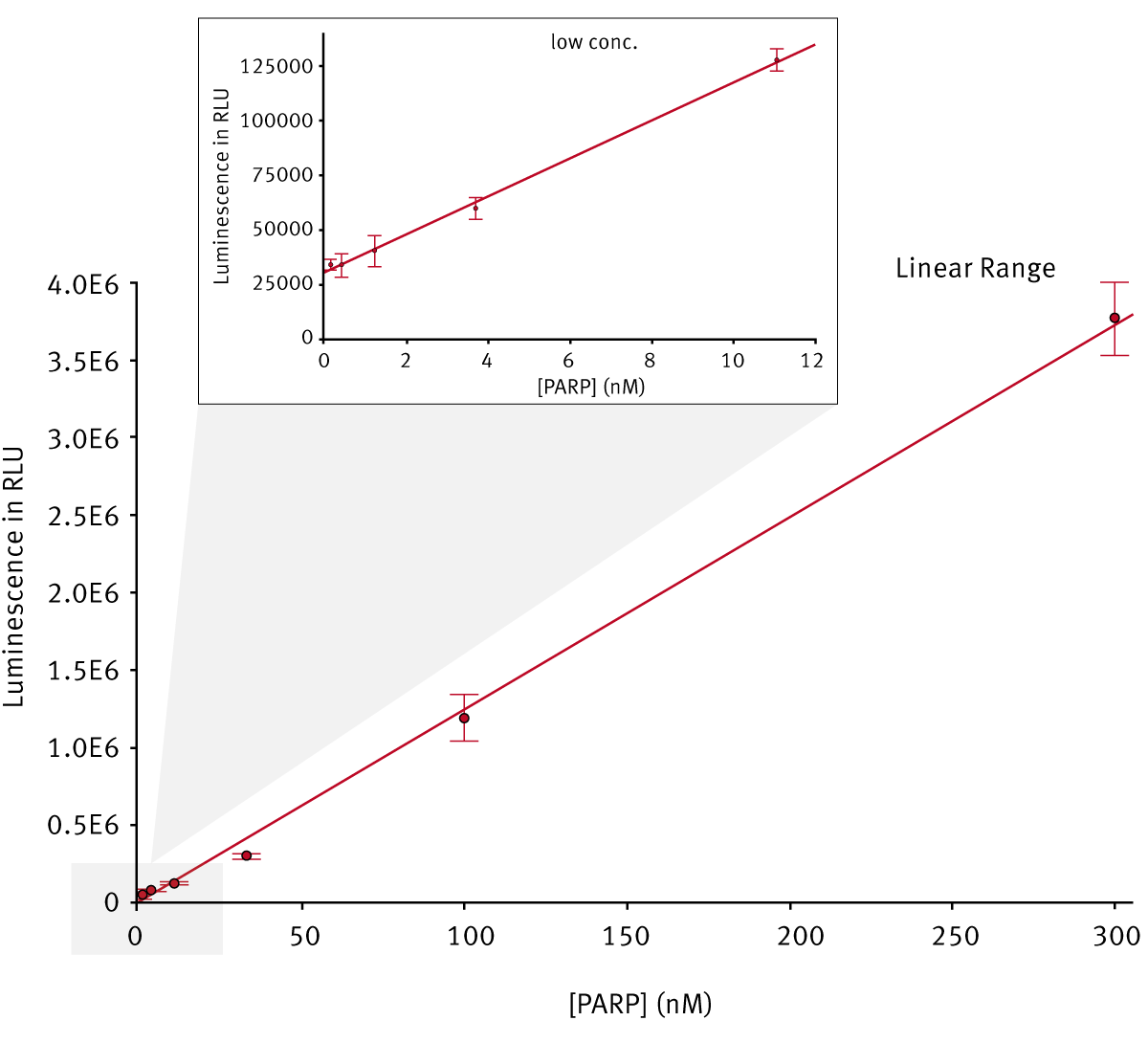 The kinetic parameters of a PARP enzyme family member were determined using initial reaction rates. Independent experiments showed that the biotin moiety linked to the co-substrate had no influence on the reaction kinetics (results not shown).
The kinetic parameters of a PARP enzyme family member were determined using initial reaction rates. Independent experiments showed that the biotin moiety linked to the co-substrate had no influence on the reaction kinetics (results not shown).
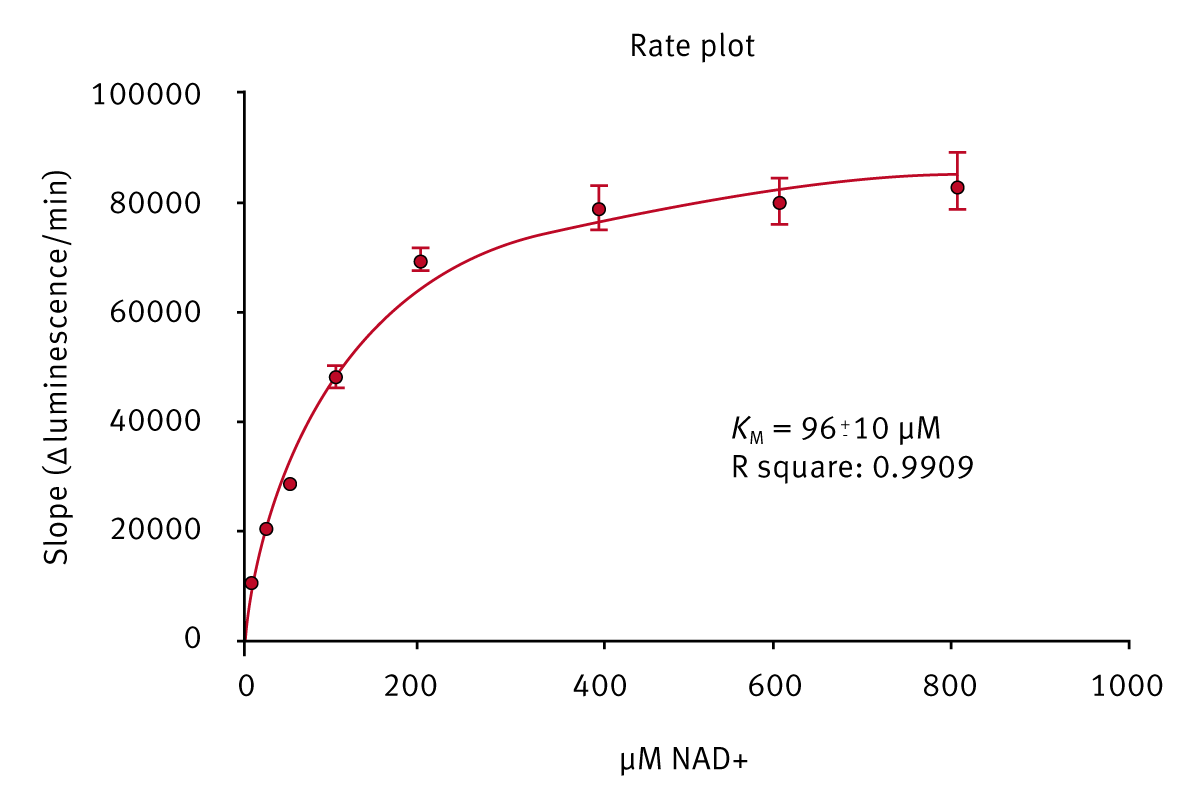
Knowledge of KM allowed the determination of inhibitor dose-response curves and experimental parameters (IC50).
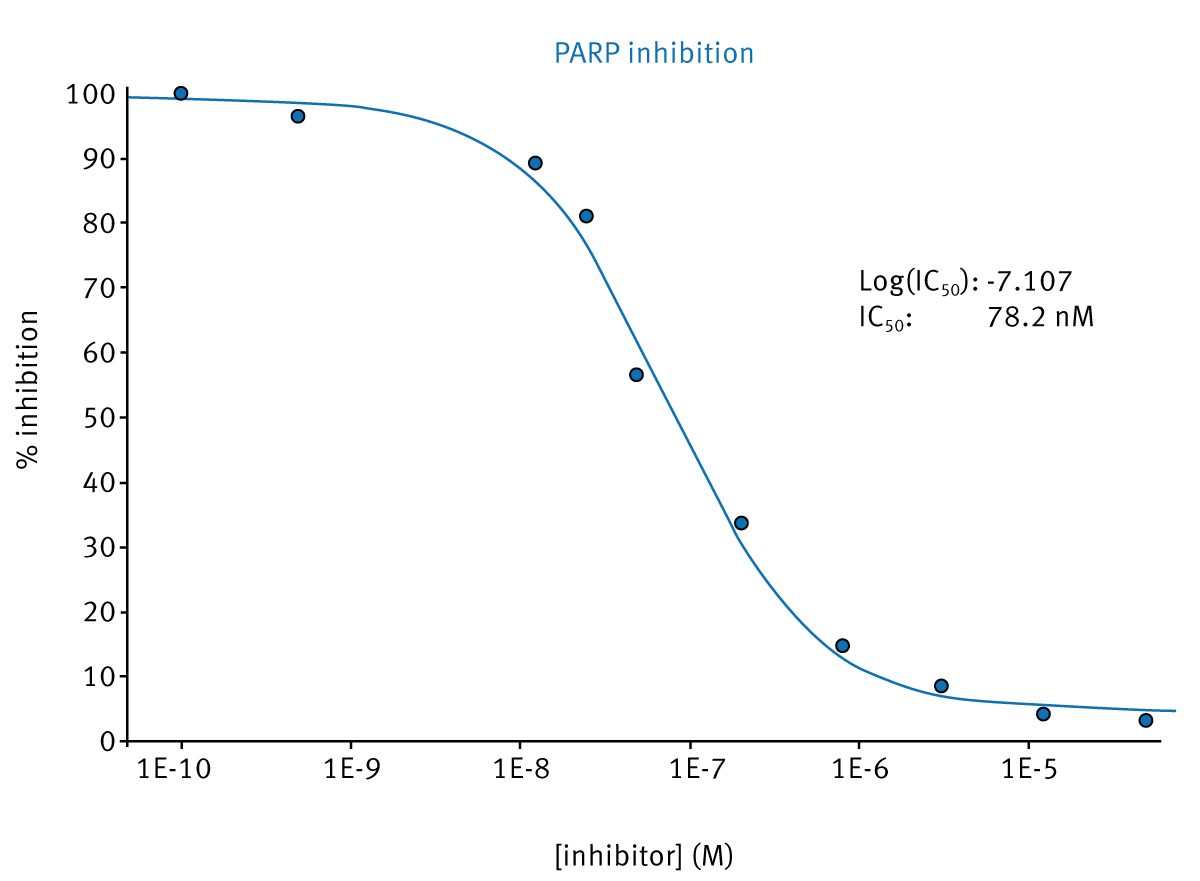
Conclusion
An ADP-ribosylation assay of PARP enzymes carried out in a BMG LABTECH microplate reader shows signal linearity over a wide range of enzyme concentrations (0.015 to 300 nM). The assay allows enzyme characterization and calculation of different parameters that are important for the development of drug like enzyme inhibitors.