Introduction
Streptococcus agalactiae (also known as group B streptococcus [GBS]) is associated with various diseases and infections in humans and animals.1 These include neonatal disease, sepsis, arthritis, pneumonia, meningitis, skin and soft tissue infections such as urinary tract infection.
Research efforts focused on the mechanisms for host colonization and virulence have mainly utilized microscopy techniques coupled with fluorescent biomarkers. Strategies for labelling GBS with fluorescent biomarkers, have so far been limited to antibody-based immunostaining methods and nonspecific protein/DNA stains.
Green fluorescent protein (GFP) expression is a common labeling method for the study of bacteria. It enables the identification of bacteria in complex samples, the monitoring of subcellular locations of bacteria in eukaryotic host cells, and the development of novel and efficient enumeration methods such as based on flow cytometry.
In a recent study1, we have reported stable expression of a green fluorescent protein mutant (GFPmut3) in GBS. This was achieved through the use of GFP-variant-expressing plasmid, pGU2664. GFPmut3 has 20x greater fluorescence intensity than GFP, allowing greater sensitivity for visualization of GBS2. Herein, we describe absorbance, fluorescence, and fluorescence polarisation intensity experiments aimed at validating and monitoring the stability of GFPmut3 expression in GBS cultures. A detailed account of microscopic analysis, effect of antimicrobials on GFPmut3 expression in GBS, and adhesion studies of GFP-expressing GBS to human epithelial cells is described in Applied and Environmental Microbiology1.
Assay Principle
Polarised fluorescence intensity
Fluorescence polarisation measures rotation of molecules in solution. High molecular weight species (like GFP) rotate slowly and when excited with plane polarised light emit a high proportion of polarised light back to the detector. Low molecular weight compounds rotate quickly and re-emit unpolarised light. The degree of polarisation (P), is defined as (Ipara - Iperp) / (Ipara+ Iperp), where Ipara is the fluorescence intensity measured parallel and Iperp is that measured perpendicular to the plane of polarization of the excitation light.
For slow rotating species in solution, the Polarisation value can be as high as 0.4 (400mP). For low molecular weight, quickly rotating molecules this value is very low (35mP or less) because Ipara-Iperp gives a very small value in the above equation due to the intensities being very nearly the same. In our study, we have made use of this property to monitor GFPmut3 fluorescence over background autofluorescence in the commonly used bacterial medium for growth of GBS, Todd-Hewitt Broth (THB). Polarisation intensities, Ipara and Iperp, were measured temporally over a 12-hour period and total fluorescence polarisation intensity was calculated by the sum Ipara+2Iperp.
Materials & Methods
- Microplates (96 well, Greiner Cat No. 655185)
- CLARIOstar with atmospheric control unit (ACU)
- Plates were sealed using gas-permeable BreatheEasy membranes (Sigma, Cat No. Z380059)
Experimental Procedure
Overnight THB cultures of GBS str 874391 were harvested by centrifugation (8,000 x g, 10 min) and washed three times with PBS (pH 7.4). Bacteria were subsequently diluted 1:100 into fresh Chemically Defined Medium (CDM) or THB (approx. 1x106 CFU per well) and 200 ul volumes of the cell suspensions were aliquoted in triplicate into wells of a 96 well plate.
Plates were sealed with breathe-easy membrane and incubated for 12h in the CLARIOstar multi-mode microplate reader, with temperature set to 37 °C and a 5 % CO2 atmosphere.
Absorbance at 600nm (OD600), Fluorescence Intensity and/or Polarised fluorescence intensity was simultaneously monitored every 15 min with agitation at 300 rpm between cycles.
Instrument Settings
|
Optic settings
|
Absorbance |
|
| Discrete wavelength | 600 nm | |
| Pathlength correction | 200 µl | |
| Fluorescence Intensity | ||
| LVF | 470-15 Auto 488.8 515-20 |
|
| Gain | 1700 | |
| Fluorescence Polarization (FP) | ||
| Filters | Ex 482-16 LP504 EM 515-30 |
|
| Target mP | 35 | |
| Gains |
A: 951; B 1113 | |
|
General settings
|
Number of flashes: |
10, 50 for FP |
|
Setting time: |
0.2 s |
|
|
Orbital averaging |
3 mm |
|
| Kinetic settings |
Number of cycles: |
74 |
|
Cycle time |
900 s |
|
|
Incubation |
37 °C |
|
|
Atmospheric Control |
CO2: 5%; O2: monitoring |
|
Results & Discussion
The GFPmut3 plasmid pGU2664 was introduced, via electroporation, into GBS 874391 to create GBS strain GU2666 (GFP+). In addition, a non-GFPmut3 vector was introduced to create strain GU2672 (GFP-).1
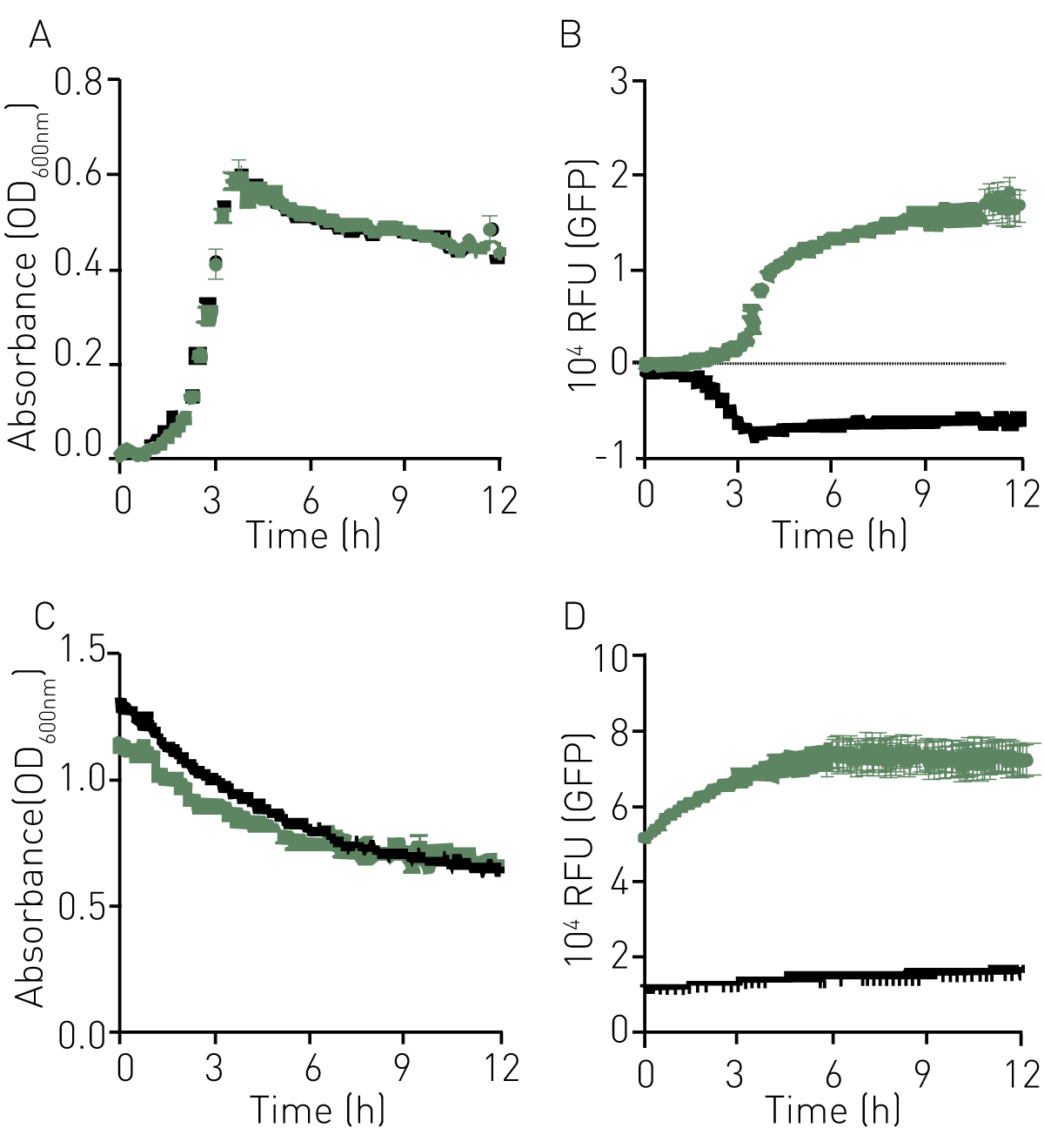
GBS containing the empty vector (GFP-), suggesting no significant metabolic burden to the GBS strain carrying the pGU2664 plasmid (Fig. 1A). In addition, there was strong GFP emission that correlated well with cell density, demonstrating expression and stability of GFPmut3 across exponential and stationary growth phases (Fig. 1B).
The fluorescence of GFPmut3 was also examined under stationary phase and low nutrient conditions. Cells were grown overnight, re-suspended in PBS, and monitored for turbidity and fluorescence over 12h, as described above. We observed stable and high-level fluorescence for these stationary phase populations with turbidity remaining stable (Fig 1, C &D).
THB is a standard complete media commonly used for growing GBS for infection experiments in animal or cell culture-based methods. We therefore also investigated the expression of GFPmut3 fluorescence during growth in THB. Interestingly, the THB media contains compounds that autofluoresce at 515nm, which interfere with the GFP signal (Fig 2B). This could be conveniently overcome by monitoring GBS growth via polarised fluorescence intensity over the 12 hour period (Fig 2C).

Conclusion
Taken together, the above results demonstrate a new method for analyzing GFPmut3 expressing GBS in experimental systems, emphasising the usefulness of the FP intensity monitoring technique in microbiology.
This study also highlights the versatility of the CLARIOstar multi-mode plate reader with ACU for microbiology. The instrument can incubate and shake samples under a controlled atmosphere (CO2 and/or O2) while acquiring data using various techniques (Fl, Lum, Abs, FP) simultaneously. In our laboratory, this has become an essential tool for investigating bacterial survival and the mechanisms of infection.
References
-
Sullivan, MJ, Ulett, GC. Applied Environmental Microbiology. (2018) 84: 1-18. DOI: 10.1128/AEM.01262-18.
-
Cormack BP et al. Gene (1996) 173: 33-38. DOI: 10.1016/0378-1119(95)00685-0 *Copyright © American Society for Microbiology, Applied, and Environmental Microbiology, 84 (18), 2018, e01262-18