Introduction
Venoms, particularly those from snakes, have long been known to affect haemostasis in prey species and human victims. Previously discovered venom activities include; thrombosis, haemolysis, hypotension, oedema, and haemorrhage. However, knowledge of the direct effects of venoms on mammalian haemoglobin are very rare in the literature and restricted to saliva from haematophagic animals. To further explore the nature of venom activity, the authors set about screening a Cardiovascular – Venom Discovery Array (T-VDACV) for effects on haemostasis, using a simple 384 well assay employing spectrometer-based microplate reader.
Materials & Methods
- Sheep blood (Harlan laboratories)
- Rabbit blood (University of Kent)
- Clear 384 well plates (Greiner)
- Cardiovascular – Venom Discovery Array (T-VDACV)
- Spectrometer-based microplate reader (BMG LABTECH)
Sheep blood or rabbit blood were lysed by addition of two volumes of distilled water and clarifi ed by centrifugation. Crude snake venoms (30 mg/ml) were derived from the Venomtech T-VDACV array.
Table 1: Snake venom was taken from the following snakes.
|
Viperidae family |
Short form |
Elapidae family |
Short form |
|
Bitis gabonica rhinoceros |
B.grh |
Naja nigricollis |
N.nig |
|
Agkistrodon contortrix contortrix |
A.cco |
Aspidelaps lubricus |
A.llu |
|
Atheris squamigera |
A.squ |
Dendroaspis polylepis |
D.pol |
|
Crotalus basiliscus |
C.bas |
Pseudechis rossignoli |
P.wei |
|
Cryptelytrops purpureomaculatus |
T.pur |
|
|
Crude venom, either neat or diluted in phosphate-buffered saline (PBS), was mixed with an equal volume of lysed rabbit blood (neat) or lysed sheep blood (diluted 1:2 in PBS) giving a final assay volume of 20 μl. Sheep blood was diluted to produce comparable absorbance readings to that of the rabbit. Plates were incubated at 30°C in the microplate reader and the absorbance spectrum (400-700 nm) read at 30 minute intervals over 16 hours.
Results & Discussion
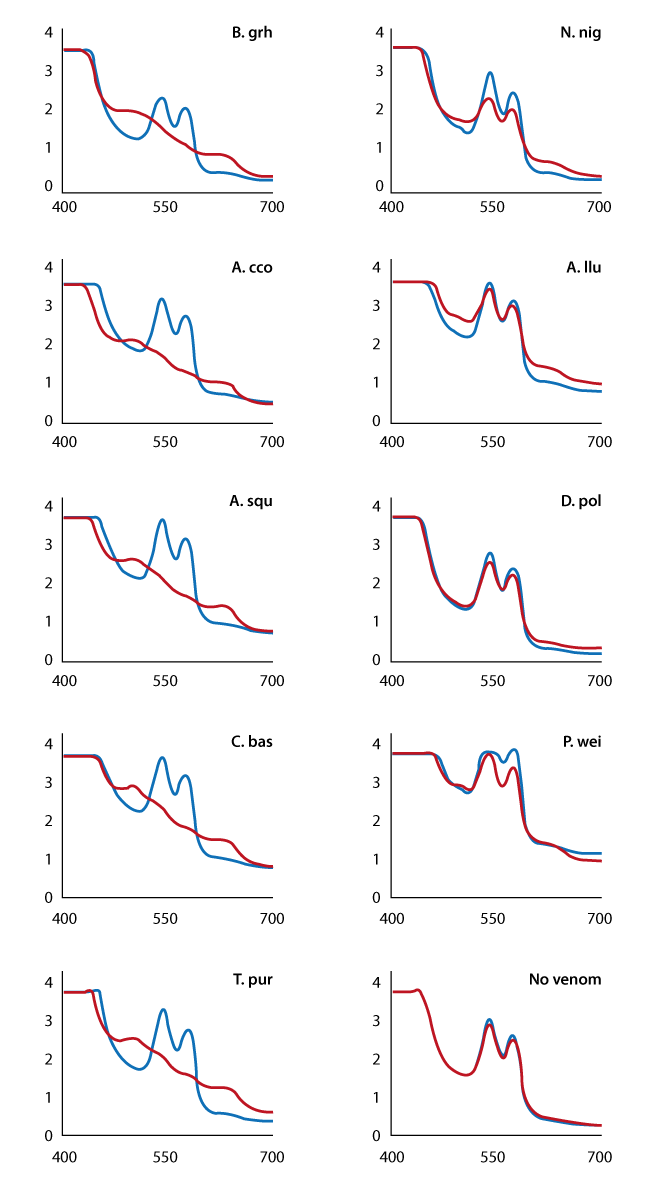
In the absence of snake venom, lysed rabbit blood exhibits absorbance peaks at 540 nm and 570 nm, characteristic of oxyhaemoglobin (HbO2) and the spectrum remained unchanged after 16 hours’ incubation at 30°C (Figure 1).
When incubated with certain snake venoms, particularly those from the Viperidae family, there was a distinct shift in the absorbance profile with a reduction of HbO2 absorbance peaks and the appearance of peaks at 500 nm and 630 nm, a profile characteristic of oxidised haemoglobin (methaemoglobin). Members of the elapidae family generally showed little activity in this assay but there was a detectable shift in the absorbance profile in the presence of the venom of the black-necked spitting cobra, Naja nigricollis. Further analysis of N. nigricollis venom, using lysed sheep blood (diluted 1:2 in PBS), demonstrated a clear time-dependent shift in the absorbance profi le from that typical of HbO2 to one more characteristic of methae-moglobin (Figure 2).
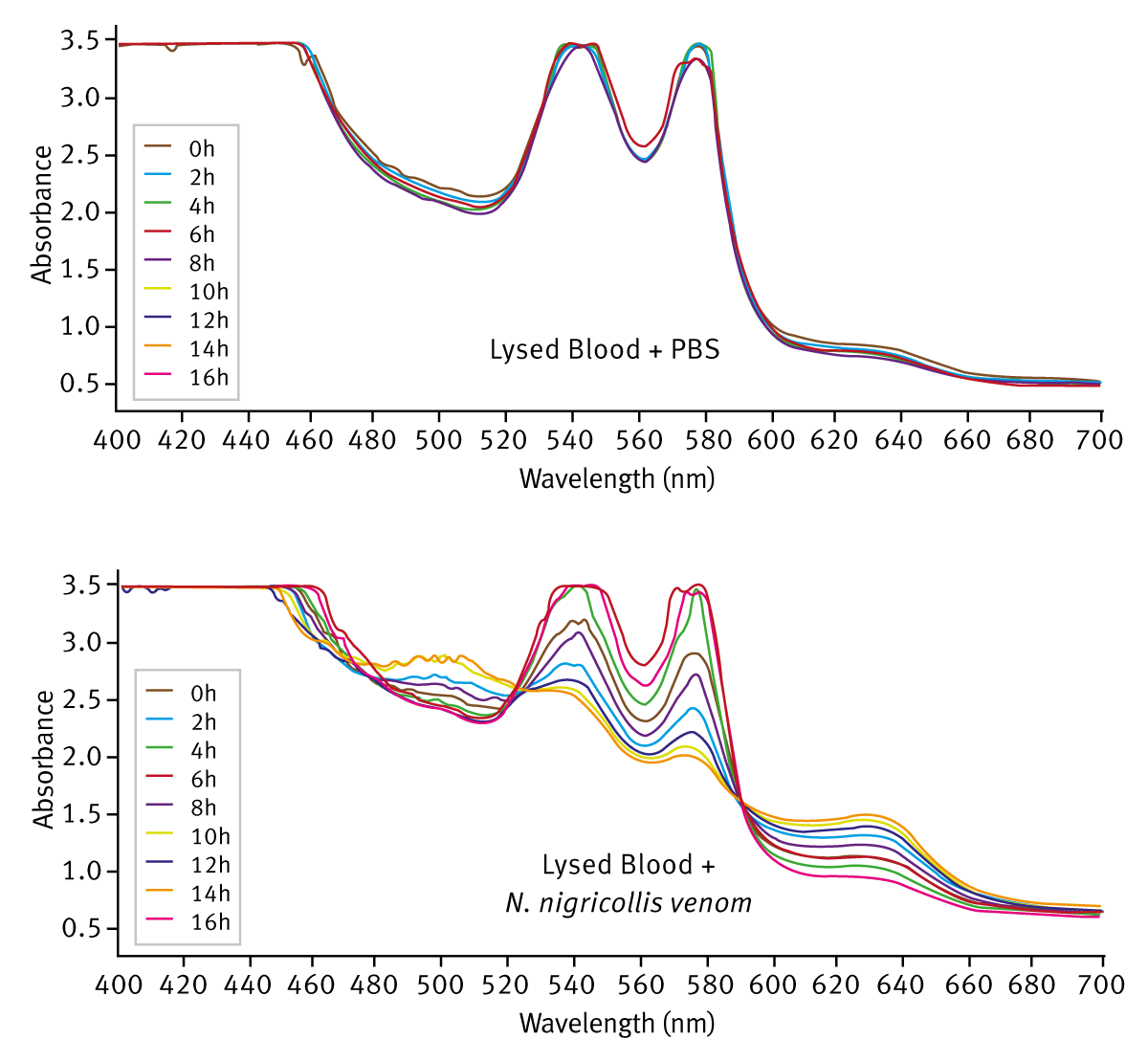
Gel-based assays showed that this spectral shift was not due to haemoglobin proteolysis (data not shown). It is also notable that oxidation of haemoglobin in the presence of N. nigricollis venom was not prevented by addition of reducing agents (dithiothreitol at 1 mM or 5 mM) whereas heat-treatment of venom (65°C or 95°C) resulted in loss of activity in this assay (data not shown.) Taken together, these data suggest that the oxidation of haemoglobin, may be enzyme-mediated and represents novel haemoglobin-modifying activity in Viperid and Elapid snake venom.
Conclusion
A simple assay employing spectral analysis of haemoglobin reveals novel haemoglobin-modifying activity in snake venom and while this effect was most pronounced in the venoms from the viperidae family, at least one elapid snake, N. nigricollis also exhibited this activity.
The microplate reader from BMG LABTECH has facilitated the design of a simple 384 well assay revealing (through wavelength scanning) novel activities in a snake venom library.