Introduction
Reactive oxygen species (ROS) are essential inter-mediates in oxidative metabolism. Nonetheless, when generated in excess, ROS can damage cells by peroxidizing lipids and disrupting structural proteins, enzymes, and nucleic acids. Excess ROS are generated during a variety of cell stresses, including ischemia/reperfusion, exposure to ionizing and ultraviolet radiation, and/or inflammation. ROS may contribute to inflammation and tissue damage.
The processes leading to ROS generation can be monitored using luminescence analysis or fluo-rescence methods. Intracellular ROS generation in cells can be investigated using 2’,7’-dichlorfluoresceindiacetate (DCFH-DA), which is an established compound to detect and quantify intracellularly produced H2O2.
The conversion of non-fluorescent DCFH-DA to the highly fluoresecent compound, 2’,7’-dichlorfluorescein (DCF), happens in several steps. First, DCFH-DA is transported across the cell membrane and deacetylated by esterases to form the non-fluorescent 2’,7’-dichlorhydrofluorescein (DCFH). This compound is trapped inside of the cells. Next, DCFH is converted to DCF through the action of peroxide, which is generated by the presence of peroxidase.
Aspergillus species are of interest in the pathogenesis of several dermatological diseases. It is uncertain whether Aspergillus itself may generate ROS and therefore actively induce tissue damage. The present study investigates whether Aspergillus species are capable of producing ROS by themselves and if there are differences between the several strains.
Materials & Methods
Six isolates of Aspergillus fumigatus (AF 65, AF 71, AF 72, AF 91, AF 210, AF 294 ) cultured 5 weeks on Sabouraud-Glucose-Agar (BAG) were investigated.
After addition of isotonic NaCl solution and centri-fugation with 100 rpm for 10 min, the blastospore concentration could be estimated by counting in CASY 1 (Schärfesystem).
These cell suspensions with concentrations of 105 to 107 cells/mL were measured on a filter-based BMG LABTECH microplate reader using 100 μL of fungal cell suspension after incubation with 100 μL DCFHDA (0.4 nM) for each single test. To eliminate LBS induced effects, polymyxin B (3 mg/mL) was added to all experimental suspensions.
Each measurement was done at least sixteen times in duplicate for calculation of the mean and the standard error of the mean.
Results & Discussion
The ability of various Aspergillus species to generate ROS was investigated.
For all fungal cells, a linear increasing fluorescence activity could be observed depending on the incubation time with DCFA-DA. By using a calibration curve, the measured fluorescence signals were converted to H2O2 concentrations and one example is given for AF 71 with different incubation periods (Fig. 1). Because of the small sized Aspergillus fumigatus (2-3 mm), detectable fluorescence was observed only at concentrations >105 cells /mL.
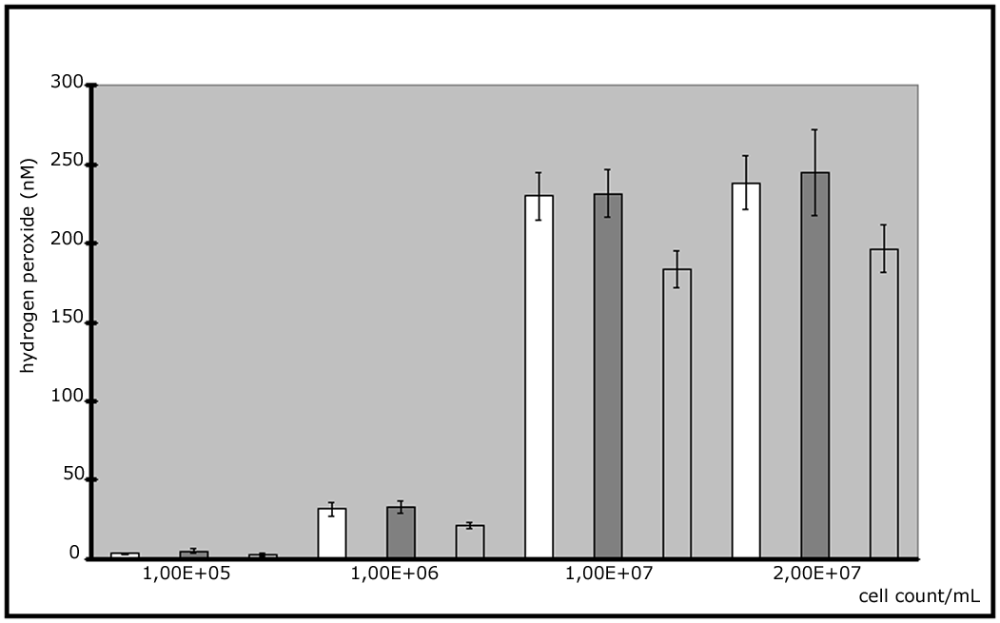
The ROS generation showed a linear and direct proportional dependence on cell numbers and the results were reproducible on 3 different days with a fixed incubation period of 2.5 hours at 37°C (Fig. 2). The highest value could be found at concentrations of 107 cells /mL.
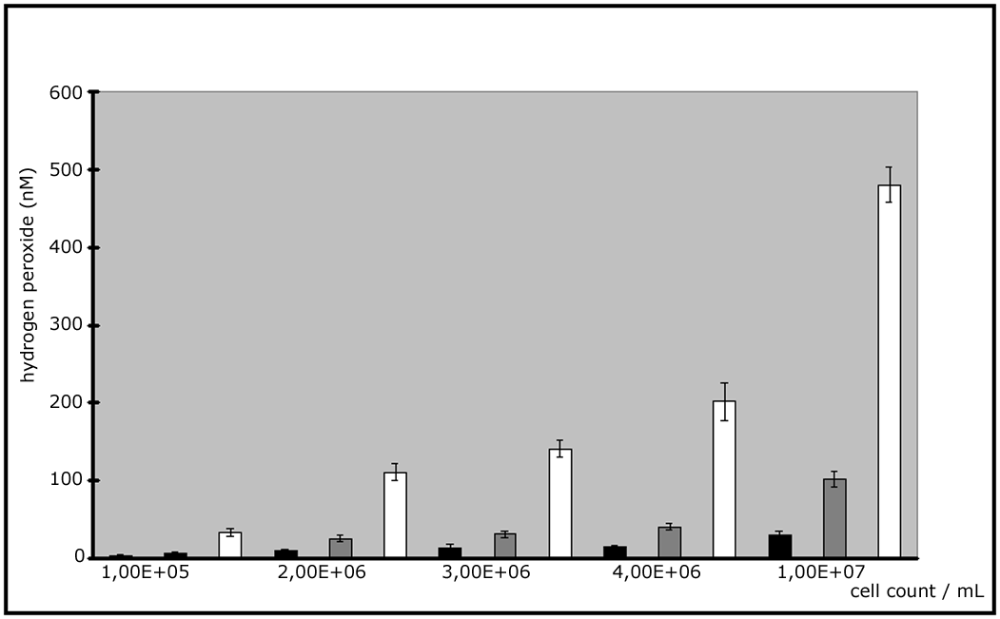
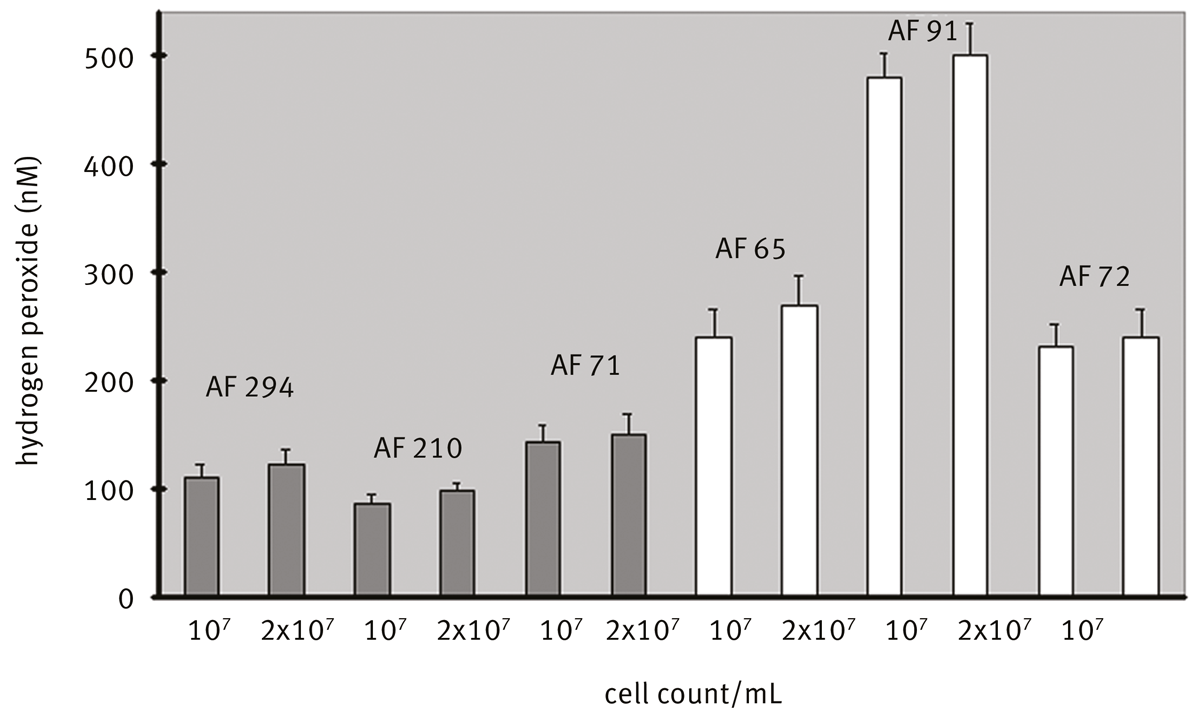
The isolates of A. fumigatus AF 91 and AF 72 are resistent against itraconazole (antifungal agent) and AF 65 is resistant to amphotericin B (antifungal agent). We investigated whether or not there are connections between the resistance and the ROS generation. Interisolate differences could be found (Fig. 3).
Conclusion
The morphological event of fungi, usually acknowledged as a major factor of virulence, is associated with increased intracellular ROS formation, which are most likely secreted. Together with phospholipases, ROS are capable of destroying the host cell membranes. This process may contribute to the invasiveness of A. fumigatus and to the inflammatory response of the host.
Clearly, the pathogenicity of Aspergillus species is a function of a multitude of parameters working together in a sequential and cooperative manner to establish infection.
It has been shown by several authors, that water as well as superoxide, hydrogen peroxide and OH-radicals can be generated in the course of the mitochondrial electron transport process. Fungi also possess the normal and the alternative pathway of electron transport. The interesting connection between the monovalent oxygen reduction and the energy conservation in isolated chloroplastes was described. The hydrogen peroxide formation at the phosphorylation points I and II is disposed by a cytochrome c oxidation process.
In this study, the method of ROS fluorescence measurement was utilized on different unstimulated Aspergillus species for the first time. There were linear correlations found between ROS levels and blastospore concentrations. A pathophysiological meaning of the released oxygen metabolites as an additional factor of virulence in the complicated system of inflammatory reactions, which was also estimated in Saccharomyces cerevisiae, is hence not to be excluded.